抽象性
后台
有充分证据表明 Arpinth感染与免疫介质疾病之间存在逆向关系多项研究显示Echioccus粮仓受感染和Hydatid细胞复合体能够抑制过敏空气道炎中的免疫响应先前的研究记录了Hydatid细胞在鼠标过敏哮喘模型中抑制异常Th2响应的能力然而,对原型学校对过敏哮喘的影响研究不足。因此,本项研究旨在评价过敏气道炎模型中的原型机体抗原效果
方法论
Ovalbumin/aumixion内注入BALB/c小鼠0至7天敏感度处理组得到原生机原型仿PBS数日,每次敏化后第14、15和16天接受1%OVA挑战剖析病理变化、支气管熔炉采集炎细胞、同化肺组织生成细胞化(IL-4、IL-5、IL-10、IL-17和IFN-GAN)和血清完全抗氧化性
结果
整体而言,通过肺同素增加二细胞素水平,将生态素录入支气管熔化液并改变病理学等方法,对原生机体抗原施用会加重过敏空气炎此外,全抗氧化容量和IFN-GAN水平经体抗原管理后下降
结论
结果显示,原型机库与OVA/alu联合管理促使BALB/c小鼠过敏气道炎加剧当前,过敏型发炎加剧的主要原因未知,需要进一步研究了解这些交互机制
图形抽象

后台
CE或Hydati一号..CE是一个大型地方卫生问题,在全世界产生相当大的经济和动物学影响,主要起因是Larval阶段Echioccus粮仓[2,3,4..成人蠕虫生长在确定宿主小肠中,在40-50天预批期[5,6..单子hydatid囊囊括原型学校,主要见于受感染中间主机的肝肺中7..
有充分证据支持指导器及其分词在预防或治疗过敏和其他炎症中作用[8,九九..浮点实验显示E.格努卢苏斯感染会通过增加插件10(IL-10)和下调血清和肺组织IL-5和IL-17A[10..特效E.格努卢苏斯一氧化二氮层评估后在严重哮喘患者中培养外围血单核细胞结果表明,薄层提取量减少了患重哮喘的病人的NO生产量11..类似抑制通过协管E.格努卢苏斯生成细胞流体敏化OVAhydatic流体内Imunodulative分子有治疗潜力抑制过敏气道炎症,通过CD4+CD25+Foxp3+T细胞大增修改免疫细胞激活和细胞平衡12..
结果确认假设E.格努卢苏斯受感染者,像在其他一些控件中一样,Hydatid细胞组件也可能对过敏空气道炎产生抑制或调控效果。然而,很少研究研究Hydatid细胞类PSC对过敏表现效果的影响
主要问题是PSC像Hydatid细胞的其他构件一样,能否抑制过敏空气道炎中的免疫响应因此,本研究旨在调查PSC体形抗原对原生过敏响应的影响
方法论
实验动物
24只母鼠BALB/c从伊朗MashhadRazi疫苗和Serum研究所获取6-8周动物无病原体寄居马沙德大学兽医学院动物设施实验室
附属点
Hydatid囊囊从Mashhad屠宰场自然感染绵羊的肝肺中收集PSC隔离式囊性流水为500x离心机g级4摄氏5分位数,隔离聚C存储到-20摄氏20分位数,供下一步使用
准备抗原
隔离PSC使用超声波实验室同质器对冰三次(每次2分2分)(UP200W26kHz)使用Bradford解析测量抗原蛋白浓度反原存储到-80°C使用PSC隔离后,从蛋白抗原中去除plioplyscharide
TritonTM X-114CAS 9036-19-5Sigma-AldrichStLouis-114最终浓度为1%v/v溶液在4摄氏度30分注入恒定扰动采样后转到37°C水池并持续扰动,孵化10分钟后200xxg级20分钟37摄氏度上半装蛋白通过管道拆分TX-114层重复TX-114辅助提取提高LPS去除效率时,重复提取程序一二三次13..安卓毒水平比原创内容下降99%完成后,含有蛋白质的上或水相移微管并存储到-20使用下相或相交相中含表面活性物和LPS被丢弃
穆林OVA诱导空气路炎模型
Hydatid囊肿急模型效果评价小鼠分为三组,每组含八小鼠如下:
- i)
负控制组(PBS组)仅在0和7天才接受PBS内注入并挑战用Nebizer使用PBS喷雾器(UltraNebTM2000德维尔比斯市曼海姆市)14、15和16天
- 二)
阳性控制组(OVA组)对OVA双注入IP敏感度Louis,MO,USA)在0天和7天用氢氧化铝(alum,Sigma-Aldrich)并用OVA喷雾器在14天、15天和16天用NUBI
- 三)
处理组(OVA+抗原组)在0和7天用二次IP注入OVA+a数天接收20微g体液提取数 OVA喷雾器挑战数日14、15和16一号)

排程过敏空气点火鼠标模型和处理协议为了诱导过敏空气路炎症,0++s+小鼠在0+7天对IP注入OVA+lum和PBSOVA+抗原组近日接受PSS仿真PBS组在0和7天收到0.5mlPBS二次敏化后,小鼠在14、15和16天接触OVA和PBS喷雾收集ABLF、肺和血清PSCs原型学校OVA系统ovalbumin阿拉姆市铝氢氧化物IP系统intraperitoneal,PBS系统磷酸盐缓冲盐ABALF系统支气管熔化液
微分细胞计数
24小时后,小鼠被麻醉,气管被暴露并割到喉角下方获取支气管熔化液支气管细胞由三度注入0.4毫升冷聚BS后肺熔化编译共收集1mlBALF并分机4摄氏4摄氏4000分5分底液悬浮100微米PBS,渗透BALF的发炎细胞总数由电磁计测定差分细胞计数时准备并染色BALF细胞粒子
评估血清总抗氧化能力
血样取自动物心脏使用胰岛素针头并转至无菌微管存储室温度后,样本离心机8000分后转微管存储到-20摄氏度血清总抗氧化功率通过使用 NaxiferTM全抗氧化能力分析包测量减肥能力估计吸附度为490纳米时使用紫外光谱计测量
肺组织历史解析
肺部解剖胸腔如下取出鼠标左肺并用于准备病理剖分组织随后固定成10%中缓冲正则解法24 h固化甲状腺后3-5微分片使用微缩并染色二叉状-eosin评估炎性细胞渗透周期酸-Schiff(PAS)染色度染色后,染色段经微量评估,病理变化评分0至4煽动性细胞渗透程度主观分类为缺漏、最小值、微值、中度或显值此外,对形形细胞元件等结构变换进行了评价,以确定是否存在发炎细胞。
测试同质肺组织中的细胞
右肺冷冻液氮并存储到-80°C检测细胞素分析日解析并称重冷冻肺,100毫克湿组织同1ml同质缓冲内含KCL(0.5M)、Tris-Cl(1M,pH9)和TritonX-100的组织同质化同质器10.0x离心g级4摄氏度为10分,超导体使用酶链免疫感应测试检测细胞素细胞素量包括IL-4、IL-5、IL-10、IL-17和插管伽马(IFN-GAN)
统计分析
所有数据均使用GreaphPad Prism软件分析(8版)。组间差异的统计意义使用单向分析和Kruskal-Wallis测试来确定P级值 < 0.05被认为重要
结果
BALF中总单元格计数
发炎细胞数计24h后OVA气溶胶挑战,见Fig2.ABLF细胞差分计结果显示小鼠接受OVA+抗原显示ABLF细胞总计数剧增,主要细胞为eosiOVA+抗原组与OVA组相比两次对PSC施用F级2 12=71.01P级< murn01>PBS小鼠群中未见渗透炎症细胞并用PBS向它敏感和挑战
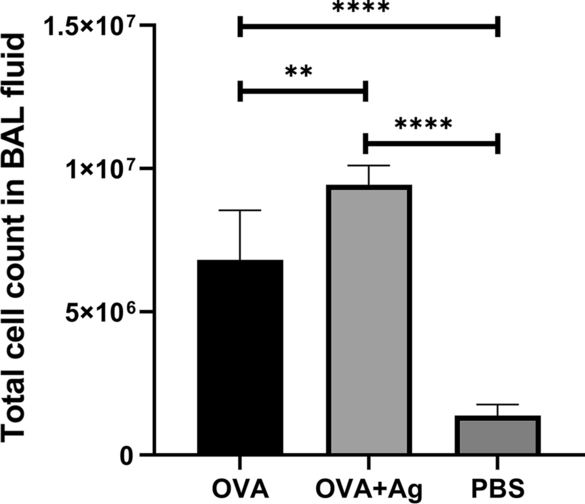
Hydatid细胞类PSC对BALB/C小鼠BALF全面炎细胞和eosi错误栏显示标准偏差P级< 0.05!单向ANOVA测试
循环分析肺同质
包括IL-4、IL-5、IL-17、IL-10和IFN-GAN3)我们的研究显示二二细胞素水平较高,包括IL-4和IL-5比照OSC管理OVA组(ANOVA:F级2 12=108.3P级<0.0001和ANOVA:F级2 12=489.0P级< 0.0001号图3ab)IL-17测量显示OVA+抗原组比OVA组高得多(ANOVA:F级2 12=142.7P级<0.0001号图3中文本不适用与OVA组相比,OVA+抗原组TH1细胞素包括IFN-GM(ANOVA:F级2 12=58.56P级<0.0001号图3公元前
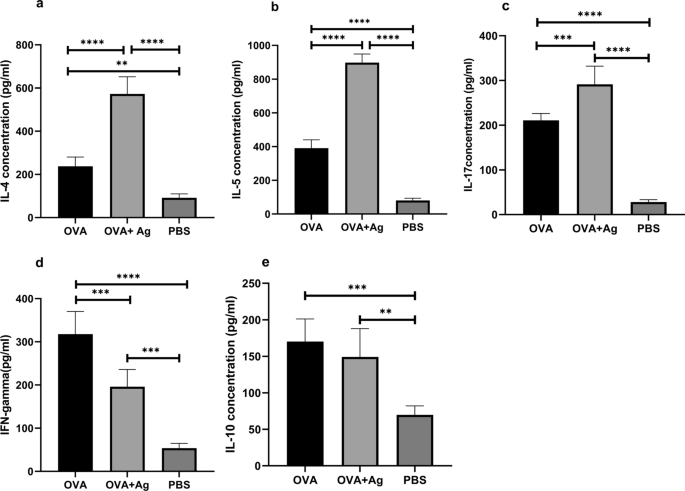
Hydatid细胞类PSC对肺组织同质中细胞级效果子宫素数由特定ELISA测试包量化ytokine响应a/IL4b/IL5C级IL17d级FNF-GMe类IL-10)使用鼠标在PSC存在或不存在时测量OVA/auce错误栏表示标准偏差P级< 0.05!单向ANOVAELISA酶关联免疫感应测试
除此以外,小鼠接受多功能类和OVA类之间IL-10水平没有显著差别(ANOVA:F级2 12=15.99P级=0.00043e)
肺病理学
OSC管理后OVA诱导小鼠的直方变化大于其他组别与OVA+抗原组和PBS组相比,指令性细胞明显渗透,主要是单核细胞,如eosiotexsχ2=12.519df=2P级=0.0024)

BALB/c小鼠OVA类肺组织骨架分解a/x20b/xx40,OVA+抗原组C级xx40d级x20和PBS组e类x20f级x20染H&EH&Eexaxilin-eosin染色
PASOVA+抗原类染色显示空气路结构变化,如与OVA和PBS群比,有标志的Goblet细胞元化和高粘合(Kruskal–Wallis:χ2=12.715df=2P级=0.0025)鼠体敏化并受PBS挑战时没有观察到病理变化

hydatid细胞PSCs对开发OVA诱导空气路炎效果a/xx40b/x20PAS系统周期酸-Schiff染色
通过减肥等离子体抗氧化力评估全反氧化能力
抗氧化能力通过血清评价发现血清总抗氧化潜能值与OVA+抗原组比明显下降OVA+PBS组(ANOVA:F级2,10=28.99P级<0.0001号图6)

hydatid细胞PSCs对血清中减肥抗氧化电量的影响以mol/l表示错误栏显示标准偏差P级< 0.05!单向ANOVA测试
讨论
近些年来,Lapinth理疗大有进展,导致引入新策略治疗免疫介质疾病。多数研究显示,拉普因感染通过确定慢性感染可引起过敏和蠕虫侵扰的症状,从而减少空气道中的eosioplia和过敏炎九九..据我们所知,这是第一份报告 说明PSC对流空气炎有充分实验证据表明,PSC和OVA/AE.格努卢苏斯抗原B(EgAGB)和Hydatid流水可加剧过敏哮喘10,11,12,14..况且,有些肠式掌上信上像Toxocarasp.,schostomes和Toxoplasmagondi寄生虫逆向关联过敏[15..
对比之下,数项实验和临床研究报告结果前后不一致研究者报告,在哮喘和其他子科疾病中,掌上型感染分子与掌上型分子有直接关联某些报告显示肠舵等Ascarislubricoides,强联想矩形,Toxocaracti并安那基斯物种可直接连接过敏型炎8..
2015年Ahumada等显示增强二比特免疫响应和IgE响应阿斯卡里斯提取作用因子推导哮喘16..2020年Bakhshani等显示口服胚胎T.Cati敏化后鸡蛋和OVA挑战加剧空气道超响应性、骨膜炎和肺炎,同时提高受鼠肺IL-5水平T.Cati[17..
PSC管理引致剧变,导致炎性细胞深入ABLF,肺同质IL-4、IL-5和IL-17水平提高,OVA+抗原组比OVA和PBS组全抗氧化能力下降
不同效果可归结为某些控件高异源性,并分物种间免分极分化,尽管这一机制的确切原因不明18号..
假设在没有自带受感染的情况下,过敏蛋白可影响宿主免疫响应前文研究显示 特殊抗原E.格努卢苏斯2细胞素解导IgE分解19号..研究结果显示IgE可帮助宿主防寄生物剂,但这一保护功能从未记录在弧线单片中20码..
特别是,抗原B(AgB)、反原5(Ag5)、循环原素(EA21)、热休克蛋白70(Hsp70)和延时因子1-Beta/delta(EF-1BB/Del带CE的人对寄生虫抗原AGB、AG5、Seineprete21号..患累进CE疾病的病人中,AGB可产生IL-4和IL-13,发光免疫病理相关TH2极化22号..显示AgB能够在大型词组中诱导反炎倍型,这有助于抑制过敏空气道生态炎14..没有细节说明AgB免疫和过敏特征所涉及的分子机制
Ortona等2002年证明E.格努卢苏斯scs和流水中存在的具体抗原在与CE关联的过敏症中发挥作用,这可能损耗或保护主机19号..
在大多数CE病人中,EgEF-1ET/QQ可偏向Th1/TH2激活向Th2极化23号..然而,大多数蛋白质不是过敏物,而仅少数抗原拥有异源特性的原因不清楚[21号..
哮喘类归为TH2疾病,与增强IgE和eosibilic炎相关,这对气道哮喘病理学至关重要24码..IL-4作为哮喘中心调试器在交换B单元IgE异型中发挥着关键作用IL-5促进eosi25码..在这次研究中,发现OVA诱导小鼠相对于OVA和PBS组施用PSC后出现增强型T2-BES响应(IL-4-IL-5)结果表明激活二类豁免还可能影响空气流的回弹性 和ABLF生物数
与高水平2细胞素一致,OVA+抗原类肺同质检测到IL-17高密度与OVA和PBS组比这表明单片机体抗原可提高17级2+抗原组并发2+++IL-17后,IL-17与2citkines并发推推推二通空路阻塞26,27号,28码..研究结果显示IL-17用Th2细胞素开发过敏哮喘
包括Treg细胞在内的其他细胞群对预防肺炎至关重要29..测量肺同质中的浓度后,OVA+抗原组IL-10水平比OVA组低,但差值无关大
IFN-GM等类型1子细胞增量因哮喘加重而增加FNF-G防止2细胞介导电感炎、航道超响应和粘合生产16..
与OVA+抗原组相比,IFN-GAN水平显著下降
FRAP解析用于确定490纳米时总抗氧化功Nadeem等2005年显示,在急性加重症患者中,等离子体总抗氧化能力下降哮喘严重加重与增强氧化压力相关减氧化性等离子阳性与炎症相关联,并起推理作用推理哮喘[30码..PSC管理后血清全抗氧化能力估值显示OVA+抗原组比OVA和PBS组显著下降先前研究也强调氧化-反氧化平衡对自然肺功能的重要性31号..然而,哮喘炎响应可能植根多路11..
历史病理学发现PSC和OVA联合管理体能抗原加气道炎与OVA和PBS组相比,OVA+抗原组中支气管上出现大量细胞支气管和支气管高密核OVA+抗原类通过PAS染色检测假设IL-13连接到气道上皮细胞中的Goblet细胞增生和脉冲制作32码并代表过敏响应的临界病理特征24码s管理OVA敏化可提高IL-13级别然而,IL-13细胞素水平在本研究的肺同质中没有得到测量。因此,不可能得出跨组间肺同质细胞素水平差异的任何结论。
结论
结果表明PSC和OVA/aulue联合管理会加剧BALB/c小鼠过敏气道炎目前,加重过敏型炎症的深层原因尚不为人知,需要进一步研究以揭示这些交互机制因此,需要进一步研究以确定过敏分子是否卷入过敏症候另一重要挑战应该强调,即PSC专用过敏可加剧炎症未来研究需要识别机制控制自导自导分子和自导自导分子对过敏型炎的加重作用
提供数据和资料
本研究期间生成或分析的所有数据都包含在本文章及其附加文件中
缩写
- PSCs:
-
原型学校
- LL:
-
分层提取
- PS:
-
Lipopolysaccharide
- PBS:
-
磷缓冲盐碱
- OVA:
-
华宾
- um:
-
铝氢氧化物
- IP注入:
-
隔离注入
- BALF:
-
条形熔化液
- H&E:
-
Hematoxylin–eosin
- PAS:
-
周期酸-Schiff
- ELISA:
-
神经联动免疫感应测试
- FRAP:
-
减肥等离子抗氧化
- EgAGB:
-
Echioccus粮仓抗原B
- AHR:
-
航道超响应性
引用
audelo HiguitaNI,BrunettiE,McCloskeyC循环电解系统JClin微生物AMSOC微生物2016年;54:518-23
Kamentzky L、CanovaSG、GuarneraEA、RosenzvitMCEchioccus粮仓:从菌层提取脱氧核糖核酸可分解肥沃非肥料囊Explasitol使用2000/95:122-7
ArmyanzasC公司,Gutierrez-CuadraM公司,FariñasMC公司Hydadisis:流行病学、临床、诊断和治疗方面Rev EspQuimioter基础2015年;28:116-24
Wen H、Vuitton L、TuxunT、LiJ、ViittonDA、ZhangW等Echinococosisce:21世纪进步Clin微生物Rev amsoc微生物201932:1-39
格塞AT流行病学和公共卫生意义评审hydadis维特美德Int2020年;2020年:1至8
周 X WangWuiFiShiCMYYYY等外细胞囊从Echioccus粮仓由病人流出hydatid细胞体积:T细胞免疫函数隔离、特征描述和评价intJ寄生虫2019;49:1029–37.
HidalgoC/GarcíaMP/StooreC/RamírezJP/MonteiroKM/HellmanU/蛋白质组分析Echioccus粮仓原型电站微信寄生素2016;218:43-5
Cruza、CooperPJ、FigueiredoCA、Alcantara-NEVSNM、RodriguesLC、BarretoML过敏和免疫学全球问题:寄生虫感染和过敏J AlvergyClinImunol2017;140:1217–28.
武兹王龙市寄生蛋白处理过敏和自动机疾病前微生物2017年8:1-13
WangH、LiJ、PuH、HasanB、MaJ、JonesMK等Echioccus粮仓增强IL-10并下调IL-5和IL-17A寄生虫向量20147:1至13
AMriMGARNAUTMTUU-BOKOVA系统生成重哮喘时一氧化二氮Echioccus粮仓.Clin遍历过敏2015年5:1
金HJ、康SA、龙TS、申MH、李KJ、朴GM等治疗效果Echioccus粮仓过敏空气道炎液Explasitol使用2019;198:63-70
TeodorowiczM,PerdijkO,VerhoekI,GoversC,SavelkoulHFJ,TangY等优化Triton X-114辅助滑动搜索法显示食品蛋白PLOS一号2017;12:1-19
席尔瓦-阿尔瓦雷斯五世、福勒AM、拉莫斯AL、北野ESSLKEchioccus粮仓抗原B绑定单细胞和大型词调控细胞对炎症的反应寄生虫向量2016年9:1-17
亚兹丹巴赫什M报寄生虫和卫生假设Clin Rev过敏Imunol2004年2月26日:15-23
Ahumada五类、GarciaE类、DennisR类、RojasMX类、RondonMA类、 PerezA类等IgE响应阿斯卡里斯兆兆兆兆兆数是哮喘风险因素Clin开发过敏2015;45:1189–200.
BakhshaniAshirvanSP MalekiMHHAHHIH评价效果Toxocaracti鼠标过敏哮喘模型感染:过敏哮喘症状加重和TH2响应类型Compimunol微生物分解2020年;71:1至23
Leonardi-BeeJ, PritchardD,BrittonJA协作气喘和当前肠寄生虫感染:系统评审和元分析AMJ回文Crit Care me2006;174:514–23.
Ortona E,VachariS,MarguttiP,DelunardoF,RiganoR,ProfumoE等名词描述Echioccus粮仓sycticenococisation患者对IgE和IgG4的异同反应ClinExpImunol2002;128:124–30.
维顿地检Echinococosisation和过敏Clin Rev过敏Imunol2004年26:93-104
菲兹西蒙斯CM 法尔科内FH DunneDWHelminth过敏性寄生虫IgE及其保护作用前名仿真20145:1-12
RiganoR,ProphumoE,BruschiF,CarulliG,AzzaraA,IoppoloS等变换人体免疫响应Echioccus粮仓抗原B 及其在逃避宿主防御中可能发挥的作用感染Imun2001年:69288-96
OrtonaE,MarguttiP,VacariS,RiganoR,ProfumoE,ButtariB等延时因子1Echioccus粮仓和过敏表现 人细胞脑电解析ClinExpImunol2001年:1225:110-6
工藤市 石松波 青井市哮喘病理学前微生物2013年4:1-16
FosterPS、MartbyS、Rosenberg高频、TayHL、HoganSP、CollisonAM等建模TH2响应和气道炎理解基本机制调节哮喘发源ImunolRev.2017;278:20-40
广西市市市市过敏空气路炎症:关键玩家超出Th2细胞路径ImunolRev.2017;278:145–61.
Al-RamliW,PrefontaineD,ChouialiF,MartinJG,OlivensteinR,LemièreC等TH17相关细胞素(IL-17A和IL-17F)严重哮喘J AlvergyClinImunol2009;123:1185–7.
Moletss, HamidQ,DavoinebF,NutkuE,TahaaR,PageN等哮喘气流中IL-17增加并诱导人类支气管纤维化生成细胞素J AlvergyClinImunol2001年:108430-8
Exclett WJ、StöltingH、Oliver RA、WalkerSA、PutturF、GregoryLG等AT细胞-Myelid IL-10轴规范2-world型哮喘模型中病原IFN-GAN依赖豁免J AlvergyClinImunol2020;145:666–78.
Nadeema、RajHG和ChabraSK哮喘剧增增加氧化压力JASTMA2005年;42:45-50
AhmadAshemMHSEINQ氧化-反氧化性不平衡关系与哮喘病人疾病增量安苏拉克医疗2012年7:226-32
卡诺什TBKIL-13诱导MUC5AC生成和goblet细胞分片抗类固醇Clin开发过敏2011;41:1747–56.
Acknowledgements
作者感谢Ferdowsi大学Mashhad为这项研究提供资金453501
供资问题
Ferdowsi大学Mashhad为该项目提供财政援助46351
作者信息
作者和附属关系
交文
HB构思思想并提供手稿反馈HB设计研究SG和SPS实地作业SG、SPS和MM进行实验室工作,SG和SPS分析数据,SG编写手稿所有作者阅读并批准最终手稿
对应作者
道德申报
道德核准并同意参赛
与小鼠操作和养殖有关的所有实验协议都符合马沙德大学动物福利委员会道德委员会制定的准则(批准ID:IR.UM.REC.1398.126)。
协议发布
不适用
竞技兴趣
作者声明他们没有竞技兴趣
附加信息
发布器注解
伟德体育在线Springer自然对发布地图和机构附属关系中的司法请求保持中立
权限和权限
开放存取文章依据创用CC4.0国际许可使用,允许使用、分享、改编、分发和复制任何介质或格式,只要你适当信任原创作者和源码提供链接并显示是否修改图片或其他第三方素材均载入文章Creative公共许可中,除非素材信用栏中另有表示素材未载入文章Creative公共许可中, 法定规则不允许使用或超出许可使用范围时, 需直接从版权持有者处获取许可 。查看许可副本访问http://creativecommons.org/licenses/by/4.0/.创用公共域免责http://creativecommons.org/publicdomain/zero/1.0/)适用于本条提供的数据,除非信用线对数据另有说明。
关于此文章
点上此文章
Ghabdian S.Parande ShirvanS.S.M.M.et al.体格抗原加重过敏哮喘Echioccus粮仓BALB/C小鼠过敏空气道炎寄生虫向量1516(2022年)。https://doi.org/10.1186/s13071-021-05125-2
接收:
接受:
发布:
多尔市:https://doi.org/10.1186/s13071-021-05125-2
关键字
- 阿斯马
- 体格产品
- 赫尔明斯理疗
- Echioccus粮仓