摘要
背景
肤色是一种公认的适应性特征,在人类身上进行了广泛的研究。了解不同人群肤色适应性的遗传基础对人类进化和医学有许多意义。
讨论
最近,在广泛的地理和时间人群中鉴定与肤色变化相关的基因方面取得了令人印象深刻的进展。在这篇综述中,我们讨论了目前已知的关于肤色变异的遗传学。我们列举了全球现代人类和古人类的几个肤色适应案例,并说明了肤色适应在不同人群中发生的原因、时间和方式。最后,我们总结了与色素沉着相关的候选位点,为进一步的进化和医学研究提供了有价值的参考。
结论
以往的研究普遍表明了肤色变化背后的复杂遗传机制,扩大了我们对人口统计学历史和自然选择在塑造人类遗传和表型多样性中的作用的理解。未来的工作需要解剖世界各地众多少数民族肤色适应的遗传结构,与主要大陆群体相比,这仍然相对模糊,并解开肤色适应的确切遗传基础。
背景
自从大约10万年前,现代人类冒险走出非洲,他们遍布各大洲,形成了各种各样的栖息地,从热带地区到北极,从低地到高地。在迁移过程中,当地环境的选择性压力(如寒冷的气候、缺氧和地方病病原体)以及随机漂移导致了人群特有的遗传变异,从而进一步影响了不同的表型,如乳糖耐受性、身高、免疫系统和代谢效率。
肤色变异是人类表型多样性最显著的例子之一。它以黑色素为主,这是一种位于表皮底部的色素沉着,由黑色素细胞产生。黑色素有两种形式,泛黑素(黄红色)和真黑素(黑棕色)。前者主要在肤色较浅的人体内积累,而后者则主要在肤色较深的人体内产生[1,2,3.,4,5].此外,黑色素颗粒的数量和大小因人而异,在决定人类肤色时,甚至比两种黑色素的比例更重要[5].其他与皮肤有关的因素,例如角蛋白,也会导致皮肤颜色的变化[6,7].
在全球人口中,肤色与纬度高度相关,从根本上说,与紫外线辐射的分布高度相关。1).靠近赤道的人群往往有深色的皮肤来抵御紫外线,因为过度暴露在紫外线下可能会降低叶酸的水平[8,9]并引致皮肤癌[10,11,12,13].高纬度地区人群的浅色皮肤是维持维生素D光合作用的基础选择,这是一个依赖紫外线的过程[14,15].
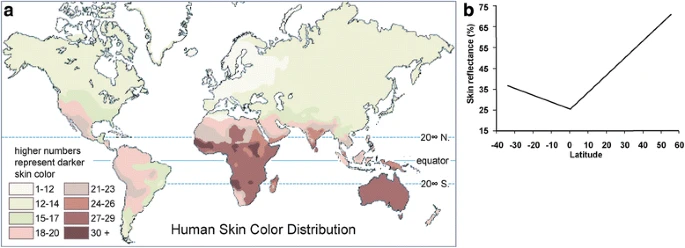
肤色与纬度的相关性(来自Barsh (2003) [5])。(一个人类肤色分布图。(b皮肤反射率与纬度的关系图
虽然紫外线被认为是人类肤色进化的驱动力,但了解选择的确切遗传机制对于重建人类进化史和阐明适应性性状的微进化至关重要。描述人类区域肤色适应的全貌将是一项挑战,因为它不仅包括被确定为处于选择状态的基因,还包括这些基因在多大程度上可以解释表型变异、基因的相互作用和联合作用,以及它们对外部环境的反应方式。在这篇文章中,我们回顾了现代人类和古人类不同种群中肤色适应的几个案例。这些案例显示了不同人群肤色适应机制的异同,并为人类进化史提供了一些见解。
现代欧亚人种的肤色适应
在欧洲,SLC24A5而且SLC45A2[16,17,18,19是两个与浅色皮肤进化有关的金色基因。SLC24A5编码NCKX5蛋白,该蛋白是跨膜蛋白家族的一员,可调节黑素小体中的钙浓度[16].该基因已被证实影响斑马鱼和小鼠的色素沉着[16,20.].特别是rs1426654的衍生等位基因SLC24A5在欧洲人中几乎是固定的,但在没有任何欧洲血统的人群中几乎没有。2) [21].大约78 kb的单倍型SLC24A5该基因与rs1426654处于连锁不平衡状态,也被确定在欧洲人中积累[22].在rs16891982中可以观察到类似的模式SLC45A2[23],据报道,它与几种物种的色素沉着有关,例如老鼠、鱼、鸟和马[24,25,26].该基因的其他变体,包括rs26722、rs2287949和rs40132,也被证明与欧洲人的颜色相关[23,27,28].在欧洲发现的另一个重要的色素相关基因是受体[29,30.,31].该基因在黑素细胞中表达,并在控制从黑素到真黑素的转换中发挥关键作用[31].色素表型与受体已在多种动物身上进行了研究[32,33,34].在受体,如rs1805007、rs1805008及rs3212357 [35,36],尽管面积较小(951 bp)。其他重要的欧洲特异基因座包括rs1393350 in酪氨酸, rs2733831 inTYRP1,rs1900758在OCA2[17,28,37,38,39].统计分析表明,这些位点的衍生等位基因频率在欧洲人身上较高,在非洲人和东亚人身上较低,这可能是欧洲人正选择的明确信号[40].
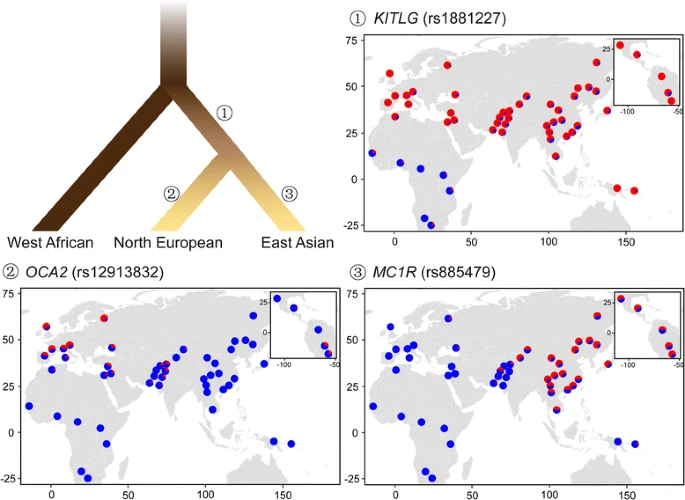
三个大陆种群中人类色素沉着的进化模型。根树显示了来自非洲、北欧和东亚的人类种群的遗传系统发育,分支的颜色大致表明了这些种群的广义皮肤色素水平(改编自McEvoy et al. (2006) [39])。据报道,现代欧亚人共同祖先中正选择的遗传位点为rs1881227KITLG,在欧洲和东亚独立进化,提示可能的趋同进化,rs12913832在OCA2和rs885479在受体,分别。等位基因频率图是用R(版本3.2.1之上,https://www.r-project.org),基于人类基因组多样性小组CEPH (HGDP)提供的53个全球人群的基因座,http://www.hagsc.org/hgdp/index.html).蓝色和红色分别表示祖先和派生的等位基因
与欧洲人发现的一长列适应基因相比,东亚人参与肤色适应的基因并没有得到很好的研究。值得注意的例子包括OCA2而且受体.每一个都有几个非同义的突变,例如rs1800414和rs74653330OCA2,和rs885479在受体[40,41,42,43],在东亚表现出较高的衍生等位基因频率,但在欧洲和非洲表现出较低的衍生等位基因频率。2).OCA2蛋白被认为是一种成熟的黑素体膜蛋白[44],在蛋白质转运到黑素体的过程中发挥潜在作用[45].rs1800414的东亚特异性变异首次在一项外显子组测序研究中报道,该研究旨在找出白化病相关的变异[46].在一项针对汉族人的关联研究中,rs1800414位点的衍生等位基因被认为有助于皮肤变白,该研究使用黑色素指数来测量个体的肤色[47].另一个非同义的变体OCA2, rs74653330,也在一项日语关联研究中被证实与色素有关[48].东亚特异性色素相关等位基因的其他例子包括rs10809814TYRP1和rs1407995DCT[40,49],两者都显示出亚洲人和非亚洲人的差异[47],以及亚洲人积极选择的强烈信号[43,49].
尽管欧洲人和东亚人在各自的地方适应下有不同的基因和变异,但一些基因衍生的等位基因在两个大陆群体中都达到了高频率。例如,KITLG表现出对非非洲人的选择性横扫[50,51,52].该基因在包括皮肤在内的多种组织中广泛表达,在器官形态发生和细胞增殖中起作用。kit配体编码KITLG被称为钢铁因子,在成人皮肤黑素细胞谱系的正常发育和维持中起着至关重要的作用[53];这已在人类、鱼类和小鼠身上得到证实[54,55,56].该基因对色素沉着的影响也在一系列关联研究中得到了证实[57,58,59,60].其中一个关键变异是rs642742,位于转录起始位点上游326 kb处KITLG.在这个变异中,非洲人的祖先等位基因频率超过90%,与欧洲人和东亚人的派生等位基因频率相当(图2)。2).在其他基因中也观察到类似的模式,例如,ASIP而且BNC2[39].
在上述结果和其他相关研究的基础上,提出了两种人类色素沉着的进化结构模型(图2)。2).一种是趋同演化模型[17,40,43,49,这表明,在某种程度上,脱色在欧洲人和东亚人是独立进化的,因为不同的基因和变异被认为可以解释这两个大陆群体的浅色皮肤和积极选择。最近的一项研究估计,欧洲特有的色素变体的选择性扫描时间大约在11000 - 19000年前,在欧洲人和亚洲人的分歧之后[61].另一种模式适用于欧洲人和东亚人共同的选择性横扫,这可能发生在原始欧亚人身上。据估计,大迁徙开始于大约3万年前,就在“走出非洲”的大迁徙之后,但早于欧洲特有的色素沉着进化[61].这两种模型的共存表明了现代人类皮肤颜色的复杂进化史。
肤色进化的复杂遗传基础的另一个线索是在单个基因中观察到的等位基因异质性,如OCA2而且受体.在这些基因中,有些等位基因是欧洲人特有的,而另一些等位基因是亚洲人特有的,尽管它们都已被证明与脱色有关。此外,OCA2提供了欧洲人和亚洲人的独立横扫以及趋同进化的证据。由于结果是从使用不同样本、数据和方法的研究中获得的,可能存在一些混杂因素导致这些不同的观察结果。然而,更重要的是,肤色是一个复杂的特征,不能简单地用单一基因或变异来解释;相反,它很可能涉及一个巨大的基因和表型网络。例如,ASIP,是欧洲血统人群中的一种适应性色素沉着基因[62,63],编码刺豚鼠信号蛋白,该蛋白会阻断受体真黑素合成对紫外线诱导的DNA损伤的响应[40].在黑色素的产生过程中,酪氨酸作为关键初始步骤的催化剂,其稳定性由TYRP1而且DCT.
此外,对皮肤色素沉着的选择扫描显示了两种不同的选择行为,分别作用于新生突变和持续变异。一些变体,以rs1805007和rs1805008为代表受体(在欧洲)和rs1800414OCA2(在亚洲),只在这些位点的正选择人群中显示出衍生等位基因,由此我们可以推测它们是现代人类定居欧洲或亚洲后出现的新突变。相比之下,一些变体,如rs3212357中的受体(在欧洲人的积极选择下),在非洲人的频率较低。不管非洲人群中可能发生的突变事件和遗传漂变,更有可能的是,该位点上的衍生等位基因在它们被青睐之前已经存在了一段时间。在藏族人的高海拔适应和一些现代人的免疫适应中,甚至在非人类物种的色素表型进化中也发现了类似的情况[56,64].
混合种群的肤色适应
混合种群,即两个先前孤立种群的杂交后代,可能为理解地理变异的遗传学提供重要的见解,原因有二。首先,祖先群体中表型差异的基因座也与信息丰富的祖先标记重叠,这使得混合群体对追溯群体历史特别有用。其次,混合群体在某些特定表型上通常存在较大的变异,这可能在控制潜在群体分层后,增加了对复杂性状/疾病相关基因的定位能力。
尽管有这些优势,混合人群在人类色素变化的研究中很少被考虑。目前在混合人群中调查色素基因的研究主要涉及具有非洲和欧洲血统的人群,如非洲裔美国人、欧洲裔非洲人和拉丁美洲人,因为他们的祖先在肤色上有很大的差异。这三个种群的祖先基因组成不同。非裔美国人从非洲血统中获得了最大的遗传贡献(~80%)[65],拉丁美洲混血儿的非洲血统比例最低(约10%)[66,67],而在欧洲非洲人中,遗传自欧洲人(~42%)和非洲人(~58%)的遗传成分是相当的[68].在拉丁美洲,有相当比例的美洲原住民血统(~45%)[66,67].此外,在个体水平上,每个祖先的比例在每个混合人口中表现出很大的差异。例如,在非裔美国人中,欧洲血统的比例从2%到98%不等[65].混种个体的肤色差异较大可能是由于他们的基因构成高度多样化,因为祖先比例与肤色之间已被观察到实质性的相关性[68,69,70].
多种已知的欧洲人色素沉着的候选基因也已通过外加剂作图确定(图2)。3.)或混合人群的关联研究。例如,酪氨酸,携带非同义替换rs1042602 (S192Y),在非裔美国人中被鉴定[69]以及来自佛得角的欧洲非洲人[68].变异的ASIP,例如rs6058017,它在全球人群中以不同的频率出现[63],也被报道与欧裔美国人的黑发和棕色眼睛有关[71]、非裔美国人[62]和巴西人[72].此外,KITLG在非裔美国人身上显示出强烈的选择性横扫信号[51],在高黑色素指数(黑皮肤)的个体中,对非洲特异性等位基因(祖先等位基因)rs642742的纯合子有显著的偏好[69].类似的情况包括rs1426654 inSLC24A5在欧洲和非洲[68]及拉丁美洲[72],并输入rs35395SLC45A2在欧洲和非洲[68].
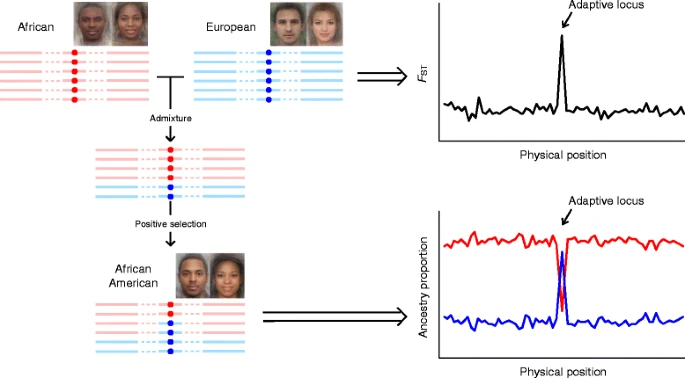
一种检测正选择的混合映射框架。非洲人、欧洲人和非裔美国人的平均面孔是从上面下载的http://www.mediadump.com/hosted-id167-average-faces-from-around-the-world.html#.WLkMU-kfU1A
然而,一些研究报告了不一致的结果。在西班牙裔人群中报道的美洲原住民血统和皮肤色素之间的相关性[73在一组波多黎各妇女中没有观察到[70].其中一个关键的单核苷酸多态性位点(SNPs)OCA2, rs1800404在分析非裔美国人和非裔美国人和非裔加勒比人的组合人群时显示出对皮肤色素沉着的显著影响,但在对非裔加勒比人样本的独立分析中不存在[69].不同人群可能存在不同的肤色变异遗传机制,但应注意数据中的详细信息,如样本量和选择用于分析混合人群的祖先人群,这可能导致有偏差的结果[67,69].
对皮肤色素沉着自然变异的遗传决定因素的鉴定也在其他混合人群中进行。一个成功的例子是对南亚后裔进行的全基因组关联研究[74的多态性SLC24A5,酪氨酸而且SLC45A2与皮肤黑色素含量有显著相关性。南亚人的浅色皮肤等位基因可能遗传自他们的欧洲祖先。75],他们最初在大约3500-4000年前随着印欧语言的扩张来到这一地区[76],随后在过去的几个世纪里被殖民。此外,中亚和东南亚是各种混合种群的家园,在肤色适应的研究中同样具有巨大的潜力。混合种群分析可以极大地丰富我们对现代人类种群中肤色变化的理解。
土著居民的肤色适应
世界各地不同地区的土著居民对人类进化史有许多影响。他们被认为是各自地区的早期定居者。尽管在某种程度上已经被周围的农学家同化,一些土著居民仍然保留着他们传统的狩猎-采集的生计,以及他们原始的身体特征——黑皮肤、矮身材和卷曲的头发。
皮肤黝黑、身材矮小、头发卷曲的狩猎采集者吸引了很多人的注意(图2)。4).这些来自中非、安达曼群岛、东南亚和大洋洲的地理位置遥远的人口(统称为Negritos或俾格米人)之间共享表型的遗传机制仍然存在争议;例如,他们是来自新石器时代之前人类底层的共同后裔,还是趋同进化的结果[77,78].迄今为止,关于这一问题的大多数遗传学研究都集中在身高上[78,79,80,81].一项研究通过分析,从皮肤色素适应的角度为趋同进化提供了线索受体美拉尼西亚人的多样性[82].这项研究表明,祖先单倍型受体在北岛美拉尼西亚人和非洲人之间并不是高度保守的,尽管这两个种群都生活在高紫外线区域,这与以前基于非常有限的样本的发现形成了对比[30.,83].此外,一个非同义多态性rs2228479在东亚人中显示出富集的衍生等位基因,但在美拉尼西亚人中与皮肤或头发色素沉着没有显著相关性。事实上,美拉尼西亚人的皮肤色素变化惊人[84],并且一致地,一些变异已被确定为区域特异性,这可以部分解释这种表型变异。一个值得注意的例子是非同义变体rs387907171TYRP1[85].它仅限于所罗门群岛和俾斯麦群岛的部分地区,可能是该地区“金发”的原因。85,86].这些结果强调了色素表型的复杂遗传结构,也强调了种群历史(例如,西南太平洋的复杂种群历史[87,88,89])可以在影响表型多样性方面发挥作用。除美拉尼西亚人外,对其他现代土著人口的皮肤色素沉着研究很少,除了一项对马来半岛的Senoi人口(土著人口)的调查,Senoi人口是Negrito(深色皮肤)和印度支那南部蒙古人种(黄棕色皮肤)的混合体,并且具有广泛的肤色谱[90].这项研究的作者发现,尽管衍生等位基因频率较低,但A111T突变(rs1426654)在该基因中存在SLC24A5与塞诺伊人的浅色皮肤显著相关,这被怀疑是蒙古人种和南亚人混合的结果。

赤道和北极地区土著居民的肤色。(一个) Bateq(黑人的一个亚群)和马来西亚半岛马来人的肤色比较。(b)来自相似纬度的因纽特人和瑞典人的肤色比较。马来人及瑞典人肖像由约书亚计划提供(http://joshuaproject.net), Bateq的肖像来自http://www.businessinsider.my/,因纽特人的肖像来自http://www.arcticphoto.co.uk/
另一个关于人类肤色适应的有趣问题来自北极人。因纽特人,在遥远的东北亚和美洲亚北极地区,尽管他们生活在遥远的北纬,但他们的皮肤是黄褐色的,不像生活在同一纬度的其他人群,如瑞典人和芬兰人(图2)。4 b).这使得因纽特人的肤色与纬度相关的分布是一个例外。一个可能的原因是深色的皮肤可以保护因纽特人免受严重的紫外线照射,因为冬季白昼时间长,雪地上的紫外线反射也很高。虽然深色皮肤不利于维生素D的产生,但包括维生素D在内的大量维生素可以从他们的饮食中得到补偿[91,92].另一个原因可能是因纽特人古代东亚血统的奠基者效应,他们从近5000年前就开始居住在北极地区,比欧洲血统产生更高的黑色素。然而,很少有遗传学研究来确定北极人群深色皮肤的遗传基础。
古人类的肤色适应性
现代人的深色皮肤大约形成于120万年前,这是由于从类人猿分化出来后体毛的减少,可能是为了防止紫外线引起的伤害[13,93,94,95,96].那么,现代欧亚人是什么时候开始脱色的呢?上述对肤色适应性的研究均基于现代群体遗传数据,可能存在群体人口统计学历史导致的有限时间分辨率,以及对长期变异作用的选择不敏感[97].古代DNA分析技术的出现使得直接观察进化过程成为可能,从而有助于我们理解这一关键问题。
一项对欧亚大陆西部(公元前6500-300年)的安纳托利亚新石器时代农民基因组的研究表明,浅色的皮肤至少在6500-4000年前就已经进化了,他们可能是第一批欧洲农民的来源人群。98].在现代欧亚人身上发现的几个流行基因,例如,SLC45A2,GRM5而且HERC2/OCA2在这些古代样本中显示出强烈的选择信号。这一结论得到了另一项基于新石器时代(6500-5000 BP)和青铜时代(5000-4000 BP)样本的研究的支持,这些样本代表了早期欧洲农民或晚期中欧狩猎采集者[99].史前欧亚大陆皮肤脱色的一个可能的动机是农业,这导致了从富含维生素d的狩猎采集饮食到缺乏维生素d的农业饮食的转变,同时在高纬度地区叶酸缺乏的危险增加了[14,One hundred.].此外,选择压力在启动肤色适应后还持续了很长一段时间,一些祖先色素等位基因在中石器时代的欧洲人(7000 BP)中被发现,一些古代欧亚人在选择下的适应性等位基因在现代人类中仍在进化[98,99,101].
最近对古人类的研究(例如,尼安德特人,一个生活在欧亚大陆约40万至28000年前的已灭绝的古人类群体[102)进一步提高了我们对现代人类肤色进化的理解。尼安德特人在大约6 - 5万年前在中东遇到了现代人,并贡献了现代人类基因组的约1-4% [103,104,105].在现代欧亚混血儿的尼安德特人的渗入单倍型中发现了一些色素相关基因,如POU2F3,BNC2而且受体[106,107].具体来说,据报道,渐渗等位基因导致浅色皮肤,这表明人类肤色适应的“适应性渐渗”策略。与皮肤表型相关的其他渐渗基因包括HYAL这些基因与细胞对紫外线的反应有关,在东亚人中存在强烈的正选择[108],以及参与角蛋白丝形成的细胞[109].虽然这些基因不是皮肤色素沉着的直接决定因素,但它们和那些与色素沉着相关的基因一样,可能帮助现代人适应非非洲环境。
当得出适应性渗透的结论时,我们实际上是在声称尼安德特人可能是浅肤色的。这一推断只是基于在现有的现代人类群体中发现的一些色素相关基因或等位基因,因为尼安德特人和其他已灭绝物种的可见表型是不可用的。然而,当使用其他一些隐修基因作为潜在线索时,可以得到不同的结果。的派生状态受体在意大利和西班牙的尼安德特人身上发现了这种色素,但在克罗地亚的尼安德特人和丹尼索瓦人身上却没有。110],这表明古人类的肤色存在差异。此外,尼安德特人和现代欧亚人的浅色皮肤也可能是趋同进化的结果,而不是适应性渗透的结果[111].
适应性遗传假说似乎早于现代人变白的时间——远早于中石器时代晚期,尼安德特人大约在28000年前灭绝。然而,我们应该重新考虑影响古人类肤色的基因是否真的决定了现代人的肤色。即使是这样,也有可能现代人类保留了这些渐渗变异,直到它们在某些特定的强选择压力下表现出一些表型效应。因此,未来需要更多的数据资源和分析来解决这个问题。
选择系数和效应量
紫外线是现代人类从非洲迁移到高纬度地区后最明显的环境变化之一,对人类皮肤色素沉着产生了相当大的选择压力,这可以通过色素沉着相关基因的选择系数来反映。选择系数的估计很大程度上取决于所考虑的基因和方法。Beleza等人估计了几个表达位点的选择系数SLC24A5,SLC45A2,TYRP1,而且KITLG[61].例如,对的估计是0.05/0.04SLC45A20.16/0.08SLC24A5在欧洲人的一种主要的/附加的遗传模式下。同时,López等报道了SLC45A2在南欧人群中为0.01-0.02 [112].这些估计值可与直接从连续采样数据推断的选择系数相比较HERC2,SLC45A2,酪氨酸,范围为0.02-0.1 [99].估计色素沉着基因的选择系数最好是在估计其他最近选择的位点的情况下理解。的选择优势被推断为0.01-0.08临床上在欧洲血统的人群中,这种基因与乳糖酶持久性密切相关[113,114], 0.019-0.048为G6PD,一种赋予非洲人群抗疟疾能力的基因[115], 0.03-0.19为EDAR与汉族人头皮毛发厚度增加和牙齿形态改变有关[116= 0.0004 ~ 0.0023EGLN1而且EPAS1有助于西藏人高海拔适应的基因区域[117].色素沉着基因的选择系数是人类基因组中最强烈选择的基因之一,表明在非洲以外的地区,紫外线或其他一些环境变化造成了严重的选择压力。
虽然已经确定了大量的基因对皮肤颜色的变化有贡献,但它们能在多大程度上解释现代人类的皮肤颜色变化呢?是否存在一种基因或变异对肤色有主要影响?某些基因可能在决定特定人群的肤色方面起着重要作用。例如,在rs1426654 in的浅色皮肤变体SLC24A5可以解释南亚地区22-32%的黑色素指数差异[75非裔美国人和非裔加勒比人的比例为25-38% [118].此外,衍生等位基因rs642742 inKITLG可以解释一个人的皮肤变浅了6到7个黑色素单位,几乎是西非人和欧洲人总体皮肤反射率差异(30个黑色素单位)的1/5 [56].然而,基因和变异相对较多,影响较小。其中一个关键的变体OCA2, rs1800414,可以解释东亚人群中约4%的色素变化[47].在南亚,rs16891982 InSLC45A2rs1042602 in酪氨酸分别占肤色变异的3.6%和2.5%,远小于rs1426654 in的效应量SLC24A5[74].皮肤色素沉着的遗传模式遵循加性模型,至少是不完全加性模型[16,17,47,56,75].
结论
总的来说,由于强大的选择压力,人类皮肤颜色是一个高度可变和复杂的特征,并由多个遗传位点控制(总结于表1).肤色适应是一个复杂的过程,因为不同种群具有共享和独立的遗传机制,涉及大量对表型具有不同影响优势的基因。肤色适应也是一个漫长的进化过程,受到各种历史甚至史前群体遗传事件的影响。目前的研究为人类肤色变化的自然选择过程和机制提供了全面的见解。更丰富的高覆盖率全基因组序列资源和表型数据可能为进一步推测遗传结构和基因环境效应的准确模型提供机会,并促进我们对某些少数民族(如狩猎采集者和高地人)皮肤色素沉着的理解。我们相信,这些研究将极大地丰富我们对人类进化史的认识,并阐明人类复杂性状的遗传基础。
参考文献
Ito S, Wakamatsu K.人类,小鼠和其他动物真黑色素和褐黑色素的定量分析:比较综述。色素细胞res 2003; 16:523-31。
Hennessy A, Oh C, Diffey B, Wakamatsu K, Ito S, Rees J. UVB照射前后人表皮真黑素和褐黑素浓度的研究。色素细胞,2005;18:220-3。
Thody AJ, Higgins EM, Wakamatsu K, Ito S, Burchill SA, Marks JM。嗜黑素和真黑素都存在于人的表皮中。中华医学杂志,2001;29(3):344 - 344。
莱利。黑色素。中国生物化学杂志,1997;29(4):339 - 339。
Barsh GS。是什么控制了人类肤色的变化?公共科学图书馆。2003;1:e27。
Tennessen JA, Akey JM。地理上不同的人类种群间的平行适应差异。PLoS Genet, 2011;7:e1002127。
高塔姆P, Chaurasia A, Bhattacharya A, Grover R.印度基因组变异联盟,Mukerji M,等。角质化基因的种群多样性和适应性进化:环境对塑造皮肤表型的影响。摩尔。杂志。进化。2015;32:555 - 73。
何云云,王晓春,金PK,赵波,范霞。蒽与叶酸络合的红外光谱和紫外光谱研究。生物化学学报。2009;42(2):366 - 366。
Juzeniene A, Stokke KT, Thune P, Moan J.健康志愿者和牛皮癣患者紫外线照射前后叶酸状态的初步研究。中国生物医学工程学报,2010;
杨晓明,杨晓明。紫外线诱发皮肤癌的流行病学研究。中华光化学杂志,2001;23(3):344 - 344。
De Gruijl FR, Van Kranen HJ, mullender LHF。紫外线诱导的DNA损伤、修复、突变和皮肤癌的致癌途径。光化学学报,2001;23(3):319 - 319。
布伦纳M,听力VJ。黑色素对人体皮肤紫外线伤害的保护作用。光化学学报。2008;84:539-49。
皮肤癌是早期人类进化中黑色色素沉着的选择性力量吗?Proc R Soc B. 2014; 181:20132955。
贾布伦斯基NG,卓别林G.人体皮肤色素沉着适应紫外线辐射。中国科学(d辑),2010,27(2):366 - 366。
哈特PH,戈尔曼S,芬雷-琼斯JJ。紫外线辐射对免疫系统的调节:不仅仅是维生素D的影响?中华免疫杂志2011;11:584-896。
Lamason RL, Mohideen MA, Mest JR, Wong AC, Norton HL, Aros MC,等。SLC24A5,一种假定的阳离子交换剂,影响斑马鱼和人类的色素沉着。科学。2005;310:1782-36。
李志强,李志强,李志强,等。欧洲人和东亚人浅色皮肤趋同进化的基因证据。中国生物医学杂志,2007;24:710-22。
Soejima M, Tachida H, Ishida T, Sano A, Koda Y.最近在欧洲人群中人类AIM1位点阳性选择的证据。分子生物学杂志,2006;23:179-88。
姜哲RS, Askew SE, Ogborne RM, Wilson S, Ferdinando D, Dadd T,等。SLC24A5编码一种具有钾依赖性钠钙交换活性的反式高尔基网络蛋白,调节人类表皮黑素生成。中国生物医学工程学报。2008;
沃格尔P,里德RW,万斯RB,普拉特KA,特劳顿K,赖斯DS。Slc24a5 - / -小鼠的眼部白化病和低色素缺陷。兽医病理学,2008;45:264-79。
Soejima M, Koda Y.色素相关基因SLC24A5和SLC45A2两个编码snp的群体差异。国际法律医学杂志2007;121:36-9。
Canfield VA, Berg A, Peckins S, Wentzel SM, Ang KC, Oppenheimer S,等。自然选择下人类常染色体肤色位点的分子系统地理学。G3。2013; 3:2059 - 67。
Cook AL, Chen W, Thurber AE, Smit DJ, Smith AG, Bladen TG,等。基于SLC45A2/MATP, SLC24A5/NCKX5和OCA2/P位点多态性的培养人黑素细胞分析。中华医学杂志,2009;29(3):342 - 344。
Fukamachi S, Shimada A, Shima A.基因编码B的突变,一种新的转运蛋白,减少黑色素含量在medaka。Nat Genet. 2001; 28:381-5。
Mariat D, Taourit S, Guérin G. MATP基因突变导致马的乳白色被毛。《人类进化史》2003;35:19 19 - 33。
Gunnarsson U, Hellström AR, Tixier-Boichard M, Minvielle F, Bed 'hom B, Ito S,等。SLC45A2基因突变导致鸡和日本鹌鹑羽毛颜色变化。遗传学。2007;175:867 - 77。
Nakayama K, Fukamachi S, Kimura H, Koda Y, Soemantri A, Ishida T. AIM1多态性在不同肤色主要人群中的显著分布。胡姆·热内。2002;47:92-4。
Valenzuela RK, Ito S, Wakamatsu K, Brilliant MH.预测模型验证:正常人体色素变化。J法医决议2011;2:1000139。
Martínez-Cadenas C, López S, Ribas G, Flores C, García O, Sevilla A,等。同时净化选择祖先MC1R等位基因和阳性选择的黑素瘤风险等位基因V60L在南欧。生物化学学报,2013;30:2654-65。
Harding RM, Healy E, Ray AJ, Ellis NS, Flanagan N, Todd C,等。MC1R可变选择压力的证据。胡姆·热内,2000;66:1351-61。
色素沉着、黑素皮质素与红头发。中华医学杂志1999;92:125-31。
Klungland H, Vâge DI, Gomez-Raya L, Adalsteinsson S, Lien S.黑素细胞刺激激素(MSH)受体在牛毛色测定中的作用。哺乳动物基因组。1995;6:636-9。
Marklund L, Moller MJ, Sandberg K, Andersson L.黑素细胞刺激激素受体(MC1R)基因的错义突变与马的栗色有关。哺乳动物基因组。1996;7:895-9。
Takeuchi S, Suzuki H, Yabuuchi M, Takahashi S.黑素皮质素1受体可能参与调控鸡羽毛色素沉着。生物化学学报。1996;138:164 - 8。
Sturm RA。GSTP1和MC1R在黑色素瘤易感性中的作用。皮肤医学杂志,2012;166:1155-6。
Tiosano D, Audi L, Climer S, Zhang W, Templeton AR, Fernández-Cancio M,等。人类维生素D受体和肤色基因的纬向分布。G3。2016; 6:1251 - 66。
杨晓明,王晓明,韩娟,王晓明。白种人皮肤色素沉着基因的遗传变异及其与皮肤色素表型的关系。中华肿瘤学杂志,2009;
杨晓明,杨晓明,杨晓明,杨晓明。不同人种皮肤色素差异的基因分析。Hum Genet, 2007; 120:613-21。
李志强,李志强,李志强。人类色素沉着正常变异的遗传结构:一个进化的视角和模型。Hum Mol Genet, 2006; 15:176-81。
Sturm RA, Duffy DL。环境选择下的人类色素沉着基因。基因组生物学,2012;13:248。
Donnelly MP, Paschou P, Grigorenko E, Gurwitz D, Barta C, Lu RB,等。OCA2-HERC2区域和色素沉着的整体视图。胡姆·热内,2012;131:683-96。
汤浅I,梅津K, Harihara S,木户A,三吉A,斋藤N,等。两个亚洲相关编码snp在MC1R和OCA2基因中的分布。生物化学,2007;45:535-42。
Hider JL, Gittelman RM, Shah T, Edwards M, Rosenbloom A, Akey JMJ,等。探索东亚血统人群色素沉淀候选基因的积极选择特征。《生物医学杂志》2013;13:150。
Sitaram A, Piccirillo R, Palmisano I, Harper DC, Dell 'Angelica EC, Schiaffino MV,等。通过细胞质双亮氨酸基序定位到成熟黑素体是人类OCA2功能所必需的。Mol Biol Cell, 2009; 20:1464-77。
Hoyle DJ, Rodriguez-Fernandez IA, Dell'Angelica EC。OCA2与蛋白质复合物BLOC-1, BLOC-2和AP-3之间的功能相互作用,从对小鼠皮毛色素沉着的上位性分析推断。色素细胞,2011;24:275-81。
Lee ST, Nicholls RD, Jong MTC, Fukai K, Spritz RA。人P基因的组织和序列及一个新的转运蛋白家族的鉴定。基因组学。1995;26:354 - 63。
爱德华兹M, Bigham A,谭j,李s, Gozdzik A, Ross K,等。OCA2多态性His615Arg与东亚人群黑色素含量的关联:皮肤色素沉着趋同进化的进一步证据。公共科学图书馆,2010;6:e1000867。
Abe Y, Tamiya G, Nakamura T, Hozumi Y, Suzuki T.日本女性黑色素生成基因与肤色变化的关系。中华皮肤医学杂志,2013;
Alonso S, Izagirre N, Smith-Zubiaga I, Gardeazabal J, Díaz-Ramón JL, Díaz-Pérez JL,等。人类黑素基因座TYR, TYRP1和DCT选择的复杂特征。BMC进化生物学,2008;8:74。
陈建民,陈建民,陈建民,陈建民,陈建民,等。人类皮肤色素沉着相关基因的正选择特征及其对皮肤色素沉着的影响。安·胡姆·热内。2007;71:354-69。
Williamson SH, Hubisz MJ, Clark AG, Payseur BA, Bustamante CD, Nielsen R.人类基因组中最近的适应性进化定位。PLoS Genet, 2007;3:e90。
Pickrell JK, Coop G, Novembre J.世界范围内人群样本中近期积极选择的信号。Genome res 2009; 19:826-37。
kit-配体在黑素细胞发育和表皮稳态中的作用。色素细胞,2003;16:287-96。
人类皮肤和色素紊乱中黑色素细胞的自分泌和旁分泌调节。色素细胞,2004;17:96-110。
班尼特,DC, Lamoreux ML.小鼠的颜色位点——一个遗传世纪。色素细胞,2003;16:33 - 44。
Miller CT, Beleza S, Pollen AA, Schluter D, Kittles RA, Shriver MD,等。刺鱼和人类中kit配体表达的顺式调节变化和色素沉着的平行进化。细胞。2007;131:1179 - 89。
Sulem P, Gudbjartsson DF, Stacey SN, Helgason A, Rafnar T, Magnusson KP,等。欧洲人头发、眼睛和皮肤色素沉着的遗传决定因素。Nat Genet, 2007; 39:1443-52。
张敏,宋峰,梁林,南红,张杰,刘华,等。全基因组关联研究确定了几个与欧裔美国人色素沉着特征和皮肤癌风险相关的新位点。Hum Mol Genet, 2013; 22:2948-59。
范德浩,张伟,李琪,张伟,等。75个影响人类红细胞的基因位点。大自然。2012;492:369 - 75。
林BD, Mbarek H, Willemsen G, Dolan CV, Fedko IO, Abdellaoui A,等。基于荷兰双胞胎家庭样本的发色遗传力和全基因组关联研究。基因。2015;6:559 - 76。
Santos M, Mcevoy B, Alves I, Cameron E, Shriver MD, Parra EJ,等。欧洲人色素沉着变淡的时间。中国生物医学杂志,2013;30:24-35。
彭妮拉C,波克斯尔L-A,麦克唐纳SA,威廉姆斯T,西尔维斯特N,帕拉EJ,等。刺鼠信号蛋白(ASIP)基因的8818G等位基因是祖传的,与非裔美国人的深色皮肤有关。胡姆·热内,2005;116:402-6。
Zeigler-johnson C, Panossian S, Gueye SM, Jalloh M, Ofori-Adjei D, Kanetsky PA。刺鼠信号蛋白G . 8818a >G多态性频率的群体差异。色素细胞,2004;17:185-7。
陆东,娄宏,袁凯,王旭,王勇,张超,等。西藏高原人的祖先起源和遗传史。胡姆·热内。2016;99:580-94。
混合动态、自然选择与混合群体病害。荷兰:施普林格;2015.
王s, Ray N, Rojas W, Parra MV, Bedoya G, Gallo C,等。拉丁美洲混血儿基因组混合的地理格局。PLoS Genet, 2008;4:e1000037。
邓丽丽,徐松,王松。拉丁美洲种群的祖先变异与基因组自然选择足迹。科学代表2016;6:21766。
Beleza S, Johnson NA, Candille SI, Absher DM, Coram MA, Anderson TM,等。非洲-欧洲混合人群皮肤和眼睛颜色的遗传结构。PLoS Genet, 2013;9:e1003372。
Shriver MD, Parra EJ, Dios S, Bonilla C, Norton H, Jovel C,等。皮肤色素沉着,生物地理血统和外加剂映射。Hum Genet, 2003; 112:387-99。
Bonilla C, Shriver MD, Parra EJ, Jones A, Fernández JR.纽约市波多黎各妇女的祖先比例及其与皮肤色素沉着和骨密度的关系。Hum Genet, 2004; 115:57-68。
潘诺西恩,李丽娟,李丽娟,李丽娟。刺鼠信号蛋白基因多态性与人体色素沉着有关。胡姆·热内。2002;70:770-5。
de Araújo LF, de Toledo GF, Fridman C. SLC24A5和ASIP作为巴西人群的表型预测因子用于法医目的。中华医学杂志2015;17:261-6。
彭妮拉C, Parra EJ, Pfaff CL, Dios S, Marshall JA, Hamman RF,等。科罗拉多州圣路易斯谷西班牙裔的混杂及其对复杂性状基因定位的影响。Ann Hum Genet, 2004; 68:139-53。
Stokowski RP, Krishna Pant VK, Dadd T, Fereday A, Hinds DA, Jarman C,等。南亚人群皮肤色素沉着的全基因组关联研究。胡恩杰。2007;81:111 - 32。
Basu Mallick C, Iliescu FM, Möls M, Hill S, Tamang R, Chaubey G,等。南亚人和欧洲人的浅色皮肤等位基因SLC24A5具有相同的血统。PLoS Genet, 2013;9:e1003912。
Cavalli-Sforza LL, Menozzi P PA。人类基因的历史和地理普林斯顿,新泽西州普林塞特。大学出版社,1994年。
引言:重访“黑人”假说:研究东南亚人类史前史的跨学科方法。《生物学报》2013;85:7-20。
Manni F, Toupance B, Migliano AB, Romero IG, Metspalu M, Leavesley M,等。侏儒表型的进化:来自非洲、亚洲和美拉尼西亚侏儒全基因组扫描的积极选择证据。生物学报。2013;85:251-84。
Clavano-Harding AB, Ambler GR, Cowell CT, Garnett SP, al - toumah B, Coakley JC,等。菲律宾Ati Negritos的GH-IGF轴和营养状况的初步描述。临床内分泌杂志1999;51:741-7。
Dávila N, Shea BT, Omoto K, Mercado M, Misawa S, Baumann G.生长激素结合蛋白,胰岛素样生长因子- i和身材矮小的两个菲律宾侏儒种群。中华儿科内分泌杂志,2002;15(4):369 - 369。
孟迪扎巴尔,马丽丽,李志刚,李志刚。非洲矮人族表型共变基因座的适应性进化研究。Hum Genet, 2012; 131:1305-17。
诺顿HL, Werren E, Friedlaender J.北岛美拉尼西亚的MC1R多样性没有受到强烈的净化选择的限制,也不能解释该地区的色素表型变异。BMC Genet 2015;16:122。
Rana BK, Hewett-Emmett D, Jin L, Chang BHJ, Sambuughin N, Lin M,等。人黑素皮质素1受体位点高多态性。遗传学。1999;151:1547-57。
Norton HL, Friedlaender JS, Merriwether DA, Koki G, Mgone CS, Shriver MD.美拉尼西亚岛皮肤和毛发色素沉着变化。《人类科学》2006;130:254-68。
Kenny EE, Timpson NJ, Sikora M, Yee MC, Moreno-Estrada A, Eng C,等。美拉尼西亚金发是由TYRP1中的氨基酸变化引起的。科学2012;336:554 - 554。
Norton HL, Correa EA, Koki G, Friedlaender JS。在北岛美拉尼西亚,与金发颜色相关的等位基因的分布。Am J Phys human pol. 2014; 153:653-62。
李志强,李志强,李志强,等。对西南太平洋人口的基因组研究。大自然。2016;538:510-3。
刘志刚,张志刚,张志刚,等。大洋洲附近和偏远地区的桥梁:所罗门群岛的mtDNA和NRY变异。中国生物医学杂志,2012;29:545-64。
Friedlaender JS, Friedlaender FR, Reed FA, Kidd KK, Kidd JR, Chambers GK,等。太平洋岛民的遗传结构。PLoS Genet, 2008;4:e19。
杨kc, Ngu MS, Reid KP, Teh MS, Aida ZS, Koh DX,等。马来西亚半岛奥朗阿斯利部落的肤色变化。公共科学图书馆,2012;7:e42752。
Schaebel LK, Bonefeld-Jørgensen EC, Laurberg P, Vestergaard H, Andersen S.富含维生素d的海洋因纽特人饮食和炎症标志物——格陵兰岛基于人群的调查。科学通报2015;4:e40。
Kolahdooz F, Barr A, Roache C, Sheehy T, Corriveau A, Sharma S.加拿大北极地区因纽特人和因纽特育龄妇女膳食中维生素D和钙的充足性:日益受到关注。PLoS One. 2013;8:e78987。
杨晓明,张晓明。人类皮肤颜色的进化。《现代科学》2000;39:57-106。
Rogers AR, Iltis D, Wooding S. MC1R位点的遗传变异和人体毛发脱落时间。当代人类。2003:105-8。
Branda RF, Eaton JW。肤色与养分光解:一种进化假说。科学。1978;201:625-6。
知更鸟啊。人类色素沉着的生物学观点。剑桥:剑桥大学出版社;1991.
Peter BM, Huerta-Sanchez E, Nielsen R.区分持续变异和从头突变的选择性扫除。PLoS Genet, 2012;8:e1003011。
Mathieson I, Lazaridis I, Rohland N, Mallick S, Patterson N, Roodenberg SA,等。230个古代欧亚人的全基因组选择模式。大自然。2015;528:499 - 503。
王尔德S, Timpson A, Kirsanow K, Kaiser E, Kayser M, Unterländer M,等。在过去的5000年里,欧洲人的皮肤、头发和眼睛色素的积极选择的直接证据。美国国家科学院学报,2014;111:4832-7。
理查兹MP,舒廷RJ,赫奇斯RE.考古学:新石器时代开始时饮食的急剧转变。大自然。2003;425:366。
欧拉尔德I, Allentoft ME, Sánchez-Quinto F, Santpere G,蒋CWK, DeGiorgio M,等。来自7000年前中石器时代欧洲人的免疫和祖先色素等位基因。大自然。2014;507:225-8。
Finlayson C, Pacheco FG, Rodríguez-Vidal J, Fa DA, Gutierrez López JM, Santiago Pérez A,等。尼安德特人在欧洲最南端的晚期生存。大自然。2006;443:850-3。
杜兰德EY,帕特森N,赖希D,斯拉特金M.测试古老的混合物之间的密切相关的人口。生物学报。2011;28:2239-52。
傅庆林,马立克,李志强,等。尼安德特人来自罗马尼亚的早期现代人,其祖先是尼安德特人。大自然。2015;524:216-9。
付强,李华,Moorjani P, Jay F, Slepchenko SM, Bondarev AA,等。来自西伯利亚西部的4.5万年前现代人的基因组序列。大自然。2014;514:445-50。
Vernot B, Akey JM。从现代人类基因组中复活幸存的尼安德特人血统。科学。2014;343:1017-21。
丁强,胡勇,徐松,王超,李辉,张荣,等。现代人MC1R中携带功能变异Val92Met的单倍型的尼安德特起源。中国生物医学杂志(英文版);2014;
丁强,胡勇,徐松,王娟,金磊。东亚人3p21.31染色体的尼安德特人渐进是正自然选择的结果。摩尔。杂志。进化。2013;31:683 - 95。
Sankararaman S, Mallick S, Dannemann M, Prüfer K, Kelso J, Pääbo S,等。尼安德特人祖先在现代人类中的分布。大自然。2014;507:354-7。
Cerqueira CCS, Paixão-Côrtes VR, Zambra FMB, Salzano FM, Hünemeier T, Bortolini MC.通过基因组数据预测人的色素表型:从尼安德特人到詹姆斯·沃森。中国生物医学工程学报。2012;24:457 - 457。
Lalueza-Fox C, Römpler H, Caramelli D, Stäubert C, Catalano G, Hughes D,等。黑素皮质素1受体等位基因表明尼安德特人的色素沉着不同。科学。2007;318:1453-5。
López S, García Ó, Yurrebaso I, Flores C, Acosta-Herrera M,陈华,等。南欧人群SLC45A2等位基因374F上自然选择和黑色素瘤易感性之间的相互作用。公共科学图书馆,2014;9:e104367。
伊坦,鲍威尔A,博蒙特MA,伯格J,托马斯MG。乳糖耐受的起源在欧洲。科学通报。2009;5:e1000491。
王晓燕,王晓燕,王晓燕,等。遗传选择和人口统计学对乳糖酶持久性扩散的影响。科学通报。2009;4:e6369。
Tishkoff SA, Varkonyi R, Cahinhinan N, Abbes S, Argyropoulos G, destroy - bisol G,等。人类G6PD的单倍型多样性和连锁不平衡:赋予疟疾抗性的等位基因的最近起源。科学。2001;293:455 - 62。
杨晓明,王淑娟,谭杰,陈晓明,等。通过选择EDAR变体的表达在小鼠中模拟最近的人类进化。细胞。2013;152:691 - 702。
郑c, Alkorta-Aranburu G, Basnyat B, Neupane M, Witonsky DB, Pritchard JK,等。混合促进了西藏高海拔地区的基因适应。Nat Commun. 2014;5:3281。
Sturm RA。人类色素多样性的分子遗传学研究。Hum Mol Genet, 2009; 18:9-17。
郭建平,李建平,李建平,等。地理在人类适应中的作用。公共科学图书馆,2009;5:e1000500。
Flanagan N, Healy E, Ray A, Philips S, Todd C, Jackson IJ,等。黑素皮质素1受体(MC1R)基因对人体色素沉着的多效性作用。Hum Mol Genet, 2000; 9:2531-7。
Córdoba-Lanús E, Hernández-Jiménez JG, Medina-Coello C, Espinoza-Jiménez A, González A, Rodríguez-Pérez MDC,等。加那利群岛人群MC1R基因变异与散发恶性黑色素瘤易感性Arch Dermatol res 2014; 306:51-8。
杨震,钟辉,陈杰,张旭,张宏,罗旭,等。在最近的人类进化中趋同皮肤变亮的遗传机制。《Mol Biol Evol》2016;33:1177-87。
确认
感谢LetPub (www.letpub.com),以便在准备这篇手稿时提供语言上的帮助。
资金
获得国家自然科学基金资助项目(91331204和31711530221)、中国科学院战略优先研究计划(XDB13040100)和前沿科学重点研究计划(qyzj - ssw - sys009)、国家杰出青年科学基金(31525014)和上海市学术带头人计划(16XD1404700)的资助;沈祥是马普独立研究组组长,中科院青年创新促进会会员。同时感谢“万人济华”国家拔尖青年创新人才计划的支持。资助者在研究设计、数据收集和分析、发表决定或手稿准备中没有任何作用。
数据和材料的可用性
不适用。
作者的贡献
SX和LD起草了手稿。两位作者都阅读并批准了最终的手稿。
相互竞争的利益
作者宣称他们之间没有利益冲突。
伦理批准并同意参与
不适用。
出版商的注意
伟德体育在线施普林格自然对出版的地图和机构从属关系中的管辖权主张保持中立。
作者信息
作者及隶属关系
相应的作者
权利和权限
开放获取本文根据创作共用属性4.0国际许可协议(http://creativecommons.org/licenses/by/4.0/),允许在任何媒介上不受限制地使用、分发和复制,前提是您对原作者和来源给予适当的赞扬,提供到创作共用许可证的链接,并注明是否进行了更改。创作共用公共领域奉献弃权书(http://creativecommons.org/publicdomain/zero/1.0/)除另有说明外,适用于本条所提供的资料。
关于本文
引用本文
邓丽丽,徐松。人类肤色在不同人群中的适应性。Hereditas155, 1(2018)。https://doi.org/10.1186/s41065-017-0036-2
收到了:
接受:
发表:
DOI:https://doi.org/10.1186/s41065-017-0036-2
关键字
- 皮肤的颜色
- 自然选择
- 遗传适应
- 现代人类
- 陈旧的问题