摘要
鲸目动物(鲸鱼、海豚和鼠海豚)是一种哺乳动物,起源于大约5000万年前的始新世时期。尽管所有的现代鲸目动物都是专性水生哺乳动物,但早期的鲸目动物是两栖的,它们的祖先是陆地偶蹄动物,类似于小鹿。一系列中间化石记录了从陆地到水的转变,其中许多化石是在印度和巴基斯坦发现的。我们回顾了raoellid偶蹄目,以及最早的鲸类家族:pakictids, ambulocatids, remingtonocetids, protocetids和basilosaurids。我们专注于鲸类器官系统的进化,因为这些详细记录了从陆地到水的过渡。
简介
鲸、海豚和鼠海豚共同构成鲸目动物。所有现代鲸目动物都生活在水里,离开水就不能生存。尽管如此,鲸目动物是哺乳动物。与其他哺乳动物不同的是,它们会哺育幼崽;它们有三根耳骨参与声音传播(锤、砧和马镫),它们的下颌由一根骨头组成(牙齿)。
鲸目动物包括有史以来最大的动物物种之一,蓝鲸(27米长,136,000公斤),但也有一些非常小的现代代表,如小头鼠海豚(1.4米长,42公斤)。尽管体型各不相同,但所有现代鲸目动物的形状都相对相似:它们有水平的尾吸片,用于游泳;它们的前肢是鳍状肢;没有外部后肢;它们的脖子很短,身体呈流线型。
鲸目动物起源于陆地哺乳动物(Thewissen和Williams2002;Fordyce和Muizon2001).陆地哺乳动物的许多共同特征在进化过程中发生了变化,最终进化成了鲸类动物。例如,毛发是哺乳动物的特征。然而,所有的现代鲸目动物都没有毛,大概是为了减少摩擦和提高运动能力。尽管如此,有些物种的胎儿在脸上保留了一些毛发,而另一些物种的胎儿则有胡须。1).这些都是它们哺乳动物遗传的明显迹象。

泛热带海豚胎儿(LACM 94389,Stenella attenuata则),须沿主席台排列
鲸类动物陆地祖先的其他特征甚至更令人印象深刻。鲸目动物有肺,它们像其他哺乳动物一样浮到水面呼吸空气,但与鱼类不同。尽管现代鲸目动物的身体完全适应了水中的生活,但它们陆地祖先的痕迹仍然存在于鲸目动物的胚胎中:现代鲸目动物没有后肢,但它们的胚胎仍然有后肢的开始。数字2显示了四个胚胎从幼到老的排列。在早期的胚胎中,有后肢,但随着胚胎发育的继续,它们消失了。造成这些器官的基因正常表达序列的中断是造成这种情况的原因(Thewissen et al。2006).

泛热带斑点海豚的胚胎(Stenella attenuata则)覆盖大约第4至9周的胚胎发育。第一个胚胎有前肢,但没有后肢(在大多数哺乳动物中,前肢先于后肢发育)。在第二个胚胎中,后肢开始形成,但它们的发育停止,并随着胚胎的生长慢慢消失(第三和第四个胚胎)。这些胚胎不是按比例绘制的。第一个胚胎长6毫米,最后一个长17.5毫米
鲸目动物与其他海洋哺乳动物,海牛类(海牛和儒艮)和鳍足类(海豹、海狮、海象)无关。海牛类与大象的亲缘关系最密切,鳍足类与陆地食肉动物(如狗和熊)有亲缘关系。在某些方面,所有鲸目动物、海牛目动物和鳍足类动物都是相似的;它们都适应了水中的生活。例如,他们都有流线型的身体,短肢,鳍状的手和脚。在其他方面,这三个群体是不同的。例如,鲸类和海牛类几乎没有体毛,而鳍足类动物有浓密的毛发。另一方面,鲸目动物和海蛞蝓的主要推进器官是尾巴,而海狮用前肢游泳,海豹用后肢游泳。
即使在达尔文的时代,人们也知道鲸类动物有陆地祖先,但记录从陆地到水的转变的化石并不为人所知:所有的鲸化石都与现代鲸有很大的相似之处。这种情况在20世纪90年代初发生了变化,当时古生物学家们在印度和巴基斯坦发现了一系列鲸类化石中的第一批,详细记录了始新世时期(大约从5400万年前持续到3400万年前)从陆地到水的转变。鲸类的起源是化石记录中最著名的宏观进化例子之一。
系统发育:鲸类与什么有关
科学家们几乎一意识到鲸类动物有陆地祖先,就试图确定鲸类动物的近亲是什么。鲸目动物与陆地哺乳动物如此不同,以至于很难发现鲸目动物和陆地哺乳动物在解剖学上有显著的相似之处。分子生物学派上了救星,发现了鲸目动物和偶蹄动物(偶趾有蹄类动物)之间的基因相似性,而这在其他哺乳动物中是不存在的。现代偶蹄动物的代表包括猪、河马、骆驼、鹿、羊、牛和长颈鹿,在这些动物中,河马被认为是现存的鲸类动物的近亲(Nikaido et al。1999;盖茨西和奥利里2001).
然而,已知的最古老的鲸鱼化石大约有5000万年的历史,鲸鱼的近亲不太可能还活着。因此,古生物学家需要在已灭绝的偶蹄类动物中找到与鲸鱼关系最密切的偶蹄类。这发生在2007年,当时在喜马拉雅山发现了raoellids的骨架,它们被证明是鲸鱼的近亲(Thewissen et al。2007).
生物之间如何相互关联的研究被称为系统发育推断,关于系统发育的假设是由一个分支图来表示的,这是一个分支图,它把越来越密切相关的组连接成越来越紧密的分支。数字3.就是这样一张早期鲸类动物的图。鲸类类群在这个分支图上的分支与最近的工作是一致的(Thewissen等。2007;Geisler和Uhen2003;Geisler等人。2007).

图中显示鲸类动物与其陆地亲戚之间的关系。这样的图被称为分支图。在图中有更多共同分支的分类单元关系更密切
在这里,我们将概述鲸类动物起源中最重要的角色。我们将讨论它们,从raoellids开始,接着是archaeocetes,生活在始新世的古代鲸鱼,大约在5500万到3700万年前。我们将按照谱系图的顺序讨论这些问题。鲸类动物的进化在那之后继续进行,有现代代表的两个鲸亚目,齿鲸目(齿鲸,包括鼠海豚和海豚)和须鲸目(须鲸),但它们的进化在这里不讨论。最近有几篇关于齿状菌和密菌进化的综述(Fordyce和Muizon2001;比安努奇和兰蒂尼2007).
鲸鱼科:鲸类的近亲
偶蹄科是偶蹄科的一种。它包含了一小群物种,其中大多数只能从牙齿和颌骨中得知(Thewissen et al。2001,2007).头骨和骷髅以一个单一的raoellid而闻名:觅食习性(Thewissen et al。2007).Raoellids只在巴基斯坦和印度西部发现,局限于始新世中下部,大约在5500万到4500万年前。
觅食习性猫是一种大小与猫相似但形状与猫截然不同的动物。它长着长长的鼻子、长长的尾巴和细长的四肢。在每一肢的末端,有四五个脚趾,末端是蹄子,类似于鹿的蹄子。觅食习性属于偶蹄目动物,脚踝处一块骨头的形状最能说明这一点。在所有哺乳动物中,这种骨头,即黄芪或距骨(图。4),有一个铰链关节,称为滑车,在那里与胫骨(胫骨)相连。在偶蹄动物和其他偶蹄动物(以及已灭绝的鲸目动物)中,黄骨有第二个滑车,位于第一个滑车的另一端,这个第二个滑车与脚踝的剩余骨头相连。这在足部的前后方向上创造了更大的灵活性。

黄芪(踝关节骨)觅食习性(RR 224), pakicetid巴基(H-GSP 98148),现代猪(3/84)和现代鹿(2/93)。这种形状的黄芪,有近端滑车(铰链关节)和远端滑车,只出现在偶蹄动物(偶蹄动物)中。
骨头觅食习性在巴基斯坦和印度边境附近的喜马拉雅山脉高处被发现。这些化石所保存的岩石表明这些骨头是埋在一条淡水溪流里的。许多骷髅觅食习性被冲在一起,骨头混在一起。这样的位置被称为骨床,因此不可能确定哪个头骨和哪个骨头在一起。因此,骨骼觅食习性如图所示。5由许多不同个体的骨骼组成。

偶蹄动物的骨架觅食习性.这里显示的骨头来自几个人,并被地质学家A. Ranga Rao在巴基斯坦和印度边境地区发现与许多其他骨头混杂在一起(©J.G.M. Thewissen)
发现了几个头骨觅食习性(无花果。6),这对于确定它与其他哺乳动物的关系很重要。化石动物之间的系统发育可以通过对它们的形态进行编码,并使用计算机程序确定重要特征的最大相似性来确定。这样的分析结果是一个分支图,而我们的研究(Thewissen et al。2007)显示觅食习性是鲸类动物的近亲(图;3.).其中一个特征是这种关系的一个强有力的指标是耳朵骨头的形状。中耳是头骨中的一个充满空气的腔体,参与声音的传播。在大多数哺乳动物中,它的壁是由一种叫做外鼓膜的骨头构成的,就像偶蹄动物和鲸类动物一样。偶蹄动物的外鼓膜大致有半个核桃壳的形状,包围着充满空气的中耳腔。在大多数哺乳动物的耳朵周围,耳壁的厚度或多或少是恒定的,但在鲸目动物中却不是这样,它们的内壁比外壁厚得多。这种增厚的壁被称为总苞,存在于所有鲸类动物中,无论是化石的还是现代的。其他哺乳动物没有苞片,除了一种:觅食习性(无花果。7).的外鼓觅食习性有增厚的内唇,有力的指示觅食习性与鲸类动物关系密切。在过去,有总苞的外鼓膜的存在是支持鲸目中包含一个物种的主要特征,因此有时也提倡这样做觅食习性(或鲸科)被列入鲸目。虽然我们相信这种观点有一些好处,但我们倾向于反对它。在我们看来,物种级别以上的动物分类主要是科学家之间交流的工具,而不稳定的分类极大地阻碍了交流:通过包括来改变鲸类的内容觅食习性导致鲸类的不稳定。类群之间的系统发育关系最好用谱系图来表达,而分类无论如何都不能准确地反映系统发育关系(因为一个祖先物种必须包括它所有的后代物种)。Raoellidae是偶蹄动物家族已有20多年的历史;一组作者(Thewissen et al。2007它们与鲸类动物有亲缘关系,这不足以成为改变这一观点的理由。

头骨和头骨碎片的四个个体觅食习性.数字用来指单个标本。RR 209有上颚的背面和眼睛的区域;RR 210是脑壳;rr207和208是一个几乎完整的头骨,只是缺少门牙区域

图中头骨耳朵区域的细节。6208年(RR)。M3是上颌的最后一颗磨牙,下颌骨窝是颌关节。中耳在生活中充满空气,有骨壁,形状类似于果壳。标本中骨壁破碎,可见壁厚(内鼓壁)。它还暴露了中耳内部,这里充满了沉积物
骨骼的另一个惊人特征觅食习性是在它四肢的骨头里发现的。哺乳动物四肢较大的骨头通常是中空的,其中的腔内充满了骨髓。通常,在横截面上(图。8),股骨的骨髓腔(股骨)占骨骼总厚度的60%以上,而称为皮质的骨壁很薄。然而,骨头觅食习性骨髓腔更厚,因此更小,仅占骨骼的42% (Thewissen et al。2007).这一特征使骨骼沉重,而沉重的骨骼使在陆地上奔跑更加困难。这种沉重的骨骼被称为骨硬化,在涉水或水底行走的水生哺乳动物中很常见,但不包括游泳者。在河马例如,骨髓腔占股骨总厚度的55%。这有助于动物在河底行走,在那里额外的骨量可以作为压舱物。早期鲸类(巴基,Ambulocetus,见下文),骨硬化也会发生,这一比例为57%。

股骨(大腿骨)的横截面觅食习性42 (RR)。的深颜色这张照片是用偏振光显微镜和石膏滤镜拍摄的骨头化石。骨髓腔内充满沉积物(灰色的在这幅图中;http://www.neoucom.edu/audience/about/departments/anatomy/AnatFaculty/media,©J.G.M. Thewissen)
进一步证明水生栖息地为觅食习性来自它牙齿的化学成分。牙齿主要由磷酸钙组成。构成牙齿的分子中的氧气来自动物摄取的饮用水和食物。两种同位素在自然界中很常见,它们是化学性质相同,但由于原子核中有多余的中子而具有更重的原子的元素:氧-16和氧-18(其中数字反映了原子的质量)。到目前为止,氧-16是更常见的同位素(自然界中超过99%),但氧-16和氧-18之间的比例在不同的环境中有所不同,生活在水中的动物与生活在陆地上的动物有不同的比例(Roe等。1998;Clementz等人。2006).牙齿的稳定同位素研究觅食习性还认为它生活在水里(Thewissen et al。2007).
这些结果表明觅食习性是水生的,因此鲸目动物起源于水生祖先。4700万年前的偶蹄动物长得像一只小鹿,是水生动物,这似乎很奇怪,但这种行为让人想起了一种现代偶蹄动物。非洲鼠鹿(Hyemoschus aquaticus)生活在非洲中部的森林地面上,主要以水果和花卉为食。它总是待在水边,当遇到捕食者的危险时,Hyemoschus跳进水里,迅速跑到安全的地方,完全淹没在水中。一段关于这种行为的精彩视频被发布在网上www.youtube.com被称为鹰对水雪佛兰(雪佛兰是非洲鼠鹿的法语名称)。
Hyemoschus不会发生骨硬化,在水里呆的时间也相对较少。从它的形态来看,似乎觅食习性更适合水生生物Hyemoschus它可能大部分时间都生活在水中。它以水草为食是有可能的,但也有可能它来到陆地上以陆地植物为食,在某种程度上与现代河马相似。
随着鲸目动物的水生起源现在已知发生在偶蹄动物中,研究人员正在寻找raoellids的陆地亲戚。有可能这些亲戚也与河马密切相关,这将使分子和形态系统发育一致。
鲸科:最早的鲸目动物
巴基鲸目是已知的最古老的鲸目动物。尽管第一批鲸类化石是在第二次世界大战之前发现的,但这些化石是如此支离破碎,以至于它们没有被认为是鲸类。直到2001年,这些鲸鱼的骨架才被发现(Thewissen et al。2001;Nummela等。2006;Madar2007;无花果。9).Pakicetids只在巴基斯坦北部和印度西部的一些地点被发现,它们大约有5000万年的历史(始新世中期)。已知的最大的巴基类化石收藏在巴基斯坦北部的卡拉奇塔山,一个名为H-GSP Locality 62的地点。这个地方是一个骨床,散布着许多不同动物的化石。10).巴基鲸有三个属,Ichthyolestes,巴基,Nalacetus,所有这些人的头骨都在62号地点被发现。11).在一个地方有大量不相关动物的骨头,这使得不可能辨认出同一个人的所有骨头。因此,巴基鲸的骨骼是由许多不同个体的骨骼组成的,这些骨骼是根据它们的大小、它们与其他原始鲸鱼的相似性、骨骼的化学成分以及它们所在地区的相对丰度来确定的。62号地点的沉积物可以告诉我们巴基鲸的生活环境(阿斯兰和德威森)1997),其中超过60%的化石是pakictids (Thewissen et al。2001).这是一条浅浅的小溪,气候又热又干燥。每年只下几次雨,但都是倾盆大雨。河床在一年中的大部分时间里都变成了浅水池,只有在雨季才有水流动。

巴基目鲸目动物的复合骨架巴基(左),Ichthyolestes(正确的).(图片来自Thewissen et al.)2001),©自然)

H-GSP第62区化石区,巴基鲸类最丰富的地区。概述了指明特定的化石埋在哪里,还有锤提供了一个比例(图片来自Thewissen和Williams (2002),©年度审核)

四具巴基鲸目动物头骨。这四个标本都没有鼻子和牙齿,但眼眶(眼窝)被保留了下来。这表明它的鼻子与头骨的连接很弱
从外表上看,巴基鲸与现代鲸目动物完全不同。它们更像长鼻子和尾巴的狼(Thewissen et al.)。2001;www.neoucom.edu/DEPTS/ANAT/Thewissen/whale_origins/whales/Pakicetid.html).然而,巴基鱼骨架的细节却讲述了一个不同的故事;这不是一只普通的陆地掠食者。这些头骨表明,这些鲸目动物的眼眶(眼窝)位于头骨顶部,这在生活在水里但看露出水面的物体的水生动物中很常见。就像觅食习性在美国,pakicetids的四肢骨骼是骨硬化的(Madar2007),也暗示了水生栖息地,这一解释与稳定同位素证据一致(Roe等。1998;Clementz等人。2006).
Pakicetids与偶蹄动物有关,这是由分支分析(Gatesy和O’leary)所显示的2001;Geisler等人。2007;Thewissen等人。2001),并且由带有两个滑车的黄芪的存在所美丽地表明(图。4).就像鲸目动物和所有鲸目动物一样,巴基鲸目动物有一个总苞,即鼓膜骨上加厚的内唇。12).

外鼓室骨巴基现代海豚Lagenorhynchus.这种骨头像碗一样包围着中耳腔。在所有鲸目动物中,外鼓膜的内侧壁都很厚,这可以从白线,被称为总苞
大约5000万年前,在从(raoellid)偶蹄目动物进化到(pakicetid)鲸目动物的过程中,发生了显著的转变。raoellids和pakicetids都有水生适应性,在淡水溪流中涉水和行走。然而,他们的生活方式却截然不同。pakictids的齿尖(牙齿上隆起的凸起)很高,与其他齿尖之间有深谷隔开(图。13).Pakicetids也有非常不寻常的牙齿磨损,牙釉质上有很大的抛光区域,这是由牙齿与牙齿的接触引起的。这种磨损模式与吃鱼有关(O’leary和Uhen1999).此外,眼睛(眼眶)和下颌关节(下颌骨窝)后面的pakicetid头骨部分非常狭窄(图。14).这影响了咀嚼肌的附着,也影响了神经进入眼睛和鼻子的路径。这意味着pakicetids吃不同的食物,处理食物的方式也不同于raoellids,它们有不同的感觉器官。科学家们仍在试图理解这些差异到底意味着什么,但它们一定在很大程度上影响了动物的功能。从鱼类动物到巴基类动物,进而从偶蹄动物到鲸类动物的转变是一个了不起的事件,其中包括头骨及其食物加工设备的大规模重建。

牙齿觅食习性(左)与pakictids的牙齿不同(正确的)在那觅食习性有四个尖的上臼齿。pakictid上磨牙有三个尖牙。磨损面(由反射光的模式指示)可能表明pakicetids吃鱼

头骨结构的差异觅食习性(RR 207)和巴基(H-GSP 96231)。的红线表示投射在中线上的轨道(眼睛)和下颌骨窝(颌骨关节)之间的距离。的绿线表示头骨的最小宽度
综上所述,pakictids从它们的raoellid祖先那里继承了水生生活方式。眼睛的位置,四肢骨骼的骨硬化,沉积学数据和稳定同位素数据都是一致的,所有这些都表明pakicetids是浅水水域的涉水动物。
陆行鲸科:第一海洋鲸目
目前发现的陆行类化石还不到十具,但其中一具是相对完整的陆行类骨架Ambulocetus•(无花果。15)、会走会游的鲸鱼(ambulare在拉丁语中是行走的意思鲸鱼座是拉丁语中鲸鱼的意思,还有•游泳;Thewissen等人。1994,1996;Madar等人。2002).其中一个人的骨头被发现在一起,部分铰接。这个骨架包括头骨和脊柱,一个前肢和两个后肢的部分。目前只发现了少量尾椎。陆行动物的化石可分为三个属,在巴基斯坦北部和印度西北部发现了这些属的遗骸。陆行类动物化石大约有4900万年的历史(始新世中期)。

骷髅a•(H-GSP 18507)有一个很大的骨盆,可以在陆地上行走时支撑它,但尾巴和后肢是在游泳时使用的(http://www.neoucom.edu/DEPTS/ANAT/publ.html,©J.G.M. Thewissen)。的锤提供一个量表
Ambulocetus比任何pakicetid都要大得多。16),大致相当于一只大型雄性海狮的大小。这种早期鲸鱼的腿短而有力,手上有五个手指,脚上有四个脚趾。脚比手大得多。它的尾椎很结实,说明它的尾巴是肌肉发达的。就像巴基在美国,陆行类动物有骨硬化性的四肢骨骼(Madar1998).股骨的骨髓腔Ambulocetus占骨骼横截面的57%。这表明Ambulocetus生活在水中,不是快速移动的掠食者。水生生物Ambulocetus与稳定同位素数据一致(Roe et al.;1998).看起来很可能是这样Ambulocetus是一种埋伏捕食者,攻击靠近它的猎物,但不会远距离追赶。在狩猎行为中,Ambulocetus可能和现代鳄鱼很相似,而且从外表上看,Ambulocetus可能看起来像鳄鱼(http://www.neoucom.edu/DEPTS/ANAT/Thewissen/whale_origins/index.html).

骷髅Ambulocetus(前),巴基(底),©J.G.M. Thewissen
骷髅Ambulocetus有一个长鼻子,从长下颚可以看出(上颚的大部分没有保存下来)。在pakicetids中,眼睛朝上,而在Ambulocetus,它们朝向两侧,尽管它们仍然位于头骨的高处(Nummela et al。2006).这种眼位出现在水生哺乳动物中,如河马。
四肢的比例(大腿、脚和手等的相对长度)Ambulocetus与河水獭(Thewissen and Fish1997).河獭用它们的后肢和尾巴游泳,很可能Ambulocetus我也这么做了。费思(1996)讨论了哺乳动物不同游泳方式的进化(图;17).写作在发现之前Ambulocetus费什预测,现代鲸类动物的游泳模式(在背部腹侧平面移动吸虫)是在一种游泳模式之前出现的,这种游泳模式包括在长尾巴的帮助下,脚在背部腹侧扫过,类似于水獭。发现Ambulocetus表明Fish的预测可能是正确的:四肢Ambulocetus在比例上与现代河水獭相似(Thewissen和Fish1997).

Fish提出的鲸类运动进化模型(1996).鲸类动物可能跟随灰色路径在左.骷髅Ambulocetus表明它通过背部和腹部摆动移动后肢和尾巴来游泳。现代鲸目动物的尾巴在背部和腹部起伏
Ambulocetus化石只在形成于浅海的岩石中被发现,可能是在沿海沼泽或森林中。稳定同位素数据表明了这一点Ambulocetus生活在部分是淡水的环境中,这可能意味着他们在河口附近(Roe et al。1998).
鲸科:长嘴鲸目
Remingtonocetidae的最古老的代表被发现在相同的化石地点Ambulocetus但已知的最大种类的remingtonocetids来自更年轻的岩石,大约在4100万到4100万年前的印度和巴基斯坦(Gingerich et al。1997).总共有四到五个属,特点是长鼻子,占头骨长度的近三分之二。
在牙齿上,remingtonocetidds是专门的(Thewissen和Bajpai2001年,一个);它们的臼齿已经失去了pakicticds和ambulocticds的粉碎盆地。这表明remingtonocetids的饮食不同于早期的鲸类。
在属中Remingtonocetus,眼睛非常小(Thewissen和Nummela2008),但它们的耳朵很大,而且在头骨上分布得很远,这一特征增强了方向性听力。在耳朵解剖的细节上,remingtonocetids也比pakictids和ambuloctids更专门(Nummela et al.;2007).与听力相关的一个特征是下颌骨孔的大小,下颌骨孔位于牙齿后面。孔是巨大的,几乎覆盖了现代鲸目动物和雷目鲸目动物的整个下颚深度,不像巴基目动物,它更小(图。18).在所有哺乳动物中,这个孔将神经和血管输送到下牙齿和下巴,但在鲸目动物中,这并不能解释它的大小。在现代鲸类动物中,这个孔除了上面提到的神经和血管外,还携带着一大块脂肪,连接下颚和中耳,并在水下传递声音。这种垫也存在于remingtonocetids中,这表明水声传播在remingtonocetids中是有效的,这是一种明显的水生适应(Nummela等。2007,2004).

鲸类和现代齿类化石中下颌骨孔的相对高度(下颌骨孔高度除以最后一颗牙齿的下颌骨高度)
remingtonocetidds也很重要,因为它们记录了另一个主要感觉器官的进化。平衡器官位于岩骨,与外鼓膜相连。平衡器官的主要部分由三个圆形管组成,它们排列在三个平面上,彼此成直角(图2)。19).一般来说,这些管的直径,半规管,鳞片大小(Spoor和Thewissen2008),但在现代鲸类动物中,沟渠已大大减少。这种减少的原因尚不完全清楚,但有可能与颈部不活动有关(Spoor等。2002).在哺乳动物实验中,前庭结肠反射是通过刺激半规管引起的神经反射,使颈部肌肉收缩,导致头部稳定,减少身体突然运动对头部的影响。大多数现代鲸类动物的颈部都相对僵硬,如果这种反射存在的话,很可能无法稳定头部,因为颈部已经相对不动了。这可能会导致半规管过度刺激,尤其是在杂技动物中。减小根管的尺寸可以减少过度刺激的可能性,也可以限制根管的敏感性。因此,这可能会给鲸类动物表演杂技的机会。雷明鲸类和所有在桡足纲上较高的鲸类都有小的运河,但巴基鲸类有大的运河。运河没有被保存下来Ambulocetus标本。

一个轮廓椭圆和回归的身体大小(上轴,单位为10-log克)与半规管半径(on轴,单位为10毫米)的现代陆地哺乳动物(栗色)和现代鲸目动物(蓝色的).鲸类化石是巴基鲸Ichthyolestes(红色的)Remingtonocetus(橙色),即原鲸Indocetus(黄色的)和龙王蜥Dorudon(紫色的).修改自Spoor等人(2002).b现代弓头鲸内耳重建图,上面有半规管,断镫骨(黄色的)和下面的耳蜗
感觉器官的形态表明听觉对于Remingtonocetus但这一愿景并非如此。这与发现化石的岩石中的环境证据是一致的。印度Remingtonocetus可能生活在被岛屿或半岛保护的泥泞海湾里。河流可能把沉积物带进了这个海湾,水可能不透明。
remingtonocetids的后颅骨骼(Bajpai和Thewissen2000)显示这些鲸鱼腿短,但尾巴很长很有力。与Fish关于鲸目动物运动进化的假设一致,这些鲸目动物可能在水中用尾巴作为主要的推进器官,只用四肢来操纵方向,它们可能是快速游泳者,尽管半规管表明它们在陆地上的运动能力有限。现代巨型南美河水獭(Pteronura取代巴西橡胶树)有一条长尾,背部腹侧平坦,在游泳时上下摆动。这种运动方式可能是游泳的好模型Remingtonocetus.因此,remingtonocetids的外观可能类似于长鼻子的巨型水獭(www.neoucom.edu/DEPTS/ANAT/Thewissen/whale_origins/whales/Remi.html).
原鲸目:征服海洋的鲸类
最早的鲸目动物、巴基鲸目、陆行鲸目和雷明托鲸目只在印度和巴基斯坦发现。随着原鲸类的起源,鲸目动物遍布全球。原鲸类存在于亚洲、非洲、欧洲和北美的低纬度地区,它们很可能在4900万至4000万年前始新世中期在全球范围内分布(Gingerich et al.;1997;威廉姆斯1998;Geisler等人。2005).
原鲸目是一个多样化的类群,大约有15个属。其中许多都没有完整的骨架,但很明显,原兽目是一个多样化的家族,在鼻子长度和耳朵形态等特征上有很大的差异。原始鲸目动物之间的一个相似点是眼睛的位置和大小,这将它们与早期的鲸目动物区分开来(Nummela等。2006;金格里奇等人。2001年,一个).眼睛总是很大(不像remingtonocetids),面朝外侧(不像pakictids和一些remingtonocetids),并且位于远离头骨中线的厚厚的扁平头骨顶部下,称为眶上盾(不像ambuloctids, pakictids和一些remingtonocetids)。也不像早期的鲸类动物,鼻口不在鼻子的尖端(Thewissen和Bajpai2001 b).相反,它位于鼻子的后部,预示着后来鲸的气孔的形成(图。20.).现代鲸类动物的气孔位于前额两眼之间,是在水下呼吸的一种适应。

最早的鲸类动物有鼻骨(灰色的)和鼻口(黑色的)靠近鼻尖。随着鲸类越来越多地生活在水中,鼻骨退缩,鼻口迁移到头骨顶部,成为气孔(由Thewissen和Bajpai改进而来)2001 b)
头骨后面骨骼的变化很难评估,因为这些骨骼只在少数物种中已知Rodhocetus而且Artiocetus来自巴基斯坦(金格里奇等人。1994,2001 b;无花果。21),Georgiacetus来自北美(Hulbert et al.;1998;赫伯特1998).在Artiocetus而且Rodhocetus,四肢短;手有五个手指,脚有四个脚趾,脚比手大得多,有点类似Ambulocetus.这些巴基斯坦原生动物当然能够在陆地上移动,它们很可能像现代海狮一样利用土地和水:在水中狩猎,但上岸交配、生产和哺乳。由于半规管很小,陆地运动一定很慢(Spoor et al。2002).在Georgiacetus在美国,唯一已知的肢体是骨盆,它似乎没有连接到脊柱,这表明这些肢体无法支撑动物的体重。Georgiacetus可能比其他原生生物更加水生。

手和脚的原头动物Rodhocetus(由Gingerich等人修改。2001 b)
原生生物在水中的移动能力也可能有所不同。虽然早期关于原鲸骨架的报告提出了吸虫的存在(Gingerich et al。1994),现在普遍认为原生生物缺乏吸虫(Gingerich et al.)。2001 b;Buchholtz1998).游泳可能是后肢划水和尾巴前后摆动的结合。
人们对原生鲸目动物的饮食和进食形态知之甚少,但是,在那里,变异似乎也是常见的。原细胞如Babiacetus有沉重的下巴(图;23),牙齿很大,暗示饮食中包括坚硬的元素(如大鱼或其他脊椎动物的骨头)。对于其他原鱼,建议以较小的鱼类为饮食(O’leary和Uhen1999).
原鲸类通常在近岸海洋沉积物中发现,通常与碳酸盐平台(如珊瑚礁)有关(Williams1998).在印度和巴基斯坦,原鱼与remingtonocetids在同一地区被发现(Gingerich et al。1997;Bajpai和Thewissen1998;金格里奇等人。1995年,一个,b;无花果。22).前者体型更大,眼睛也更大。23),表明它们捕食不同的猎物。而印度的地方暗示了一个泥泞的泻湖丰富的植物生长(Bajpai等。2006)在一些雷明海水类中大量存在,原鱼类通常在更容易受到波浪作用的更清澈的水中沉积物中发现。

化石的挖掘,左前景,在古吉拉特邦,印度。remingtonocetids和protocetids都在印度的同一地区被发现

鲸目动物和已灭绝的海洋爬行动物(鱼龙;修改自Thewissen and Nummela (2008),©加州大学出版社)
龙王鲸科:第一个完全水生的鲸类
始新世中期晚期,大约4100万年前,一种新的鲸类出现了,这是第一个与现代鲸类相似的鲸类:Basilosauridae (Uhen1998).龙王蜥有一个鼻腔开口,向眼睛方向向后移动,形成一个通气孔,前肢有鳍状肢,尾巴末端有一个吸片,后肢很小,太小了,无法在陆地上支撑身体重量。在所有这些特征中,龙王蜥类更类似于现代鲸目动物,而不是原始鲸目动物,而且它们很可能没有离开海洋,是最早的专性鲸目动物(Kellogg1936;乌亨2004).
龙王鲸类大约有七个属,但基本上可以分为两种体型。第一个发生在属中龙王鲸它的身体像蛇一样,最大长度约为17米。龙王鲸可能是通过整个身体的弯曲运动来游泳的(布赫霍尔兹1998).龙王蜥类的第二种体型较短,短至4米。这些basilosaurids,被称为dorudontines (Uhen1998),它们有着海豚状的身体,通过尾巴的上下摆动来游动。龙王蜥在新世界和旧世界都很有名,可能生活在4500万年前到3500万年前的所有海洋中。龙王蜥的脊柱之所以如此之长,一方面是由于类群中腰椎数量的增加,另一方面也是由于每根椎骨长度的增加。
与早期的始古动物相似,但与后来的大多数鲸类动物不同,龙王蜥保留了异齿齿列,在门牙、犬齿、前磨牙和臼齿之间具有明显的形态差异(Uhen2004).这与现代鲸目动物(齿鲸目)不同,现代鲸目动物的牙齿沿着牙齿排都非常相似(这种情况被称为同质性)。早期的古细菌每半颚都有11颗牙齿(总共44颗牙齿),而龙王龙则不同,它们的上颚各少了一颗牙齿,使它们的牙齿总数达到42颗。它们的磨牙与原齿目和陆行齿目有很大的不同,在上磨牙中没有一个由三个齿尖包围的中心凹陷(O’leary和Uhen1999).因此,这些牙齿不适合碾碎食物。
在前肢方面,basilosaurids与现代鲸目动物相似,因为它们的肘关节不能单独移动,它们的手有蹼,无法识别单独的手指(Uhen2004).龙王蜥和大多数哺乳动物一样,每根手指只有三个指骨,而现代鲸目动物的指骨通常都在增加。
龙王蜥的后肢保留了早期鲸鱼和陆地哺乳动物的骨骼:股骨、胫骨、腓骨、跗骨、跖骨和趾骨(Gingerich et al.)。1990).然而,后肢的尺寸大大缩小,骨盆没有附着在脊柱上,使得后肢不适合支撑这些鲸鱼的体重。正常的现代鲸目动物没有外部后肢,尽管出生时很少有具有这种四肢的异常个体(图2)。24).在内部,所有物种都有骨盆或后肢残余物,为生殖器的肌肉提供了起源。在一些物种,骨盆,股骨和胫骨是存在的(图。25而且26).考虑到龙王龙的后肢甚至保留了足部的骨头,我们有理由认为它的一些部分是从龙王龙的身体中投射出来的。1990)提出。

在太极鲸鱼博物馆收藏的一只反常海豚的后鳍
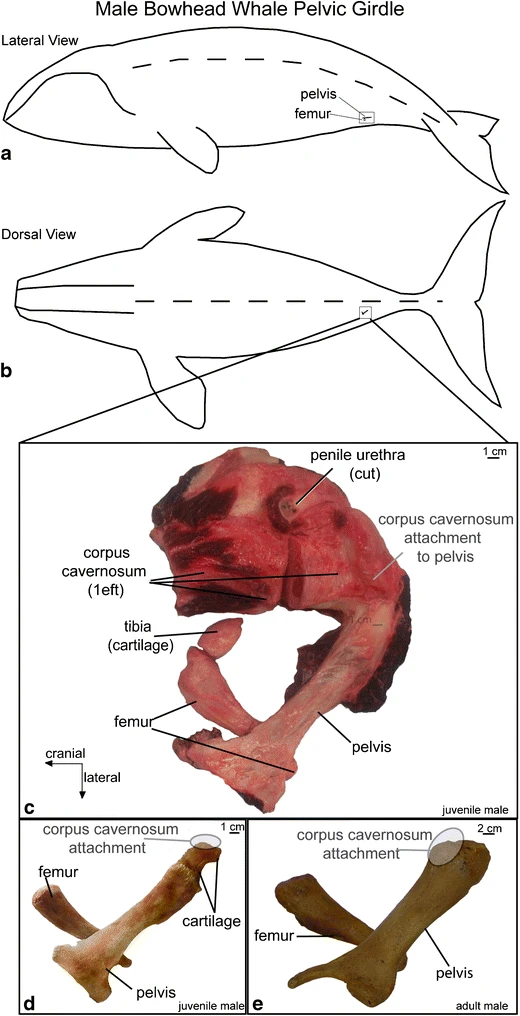

图示显示骨盆的变化觅食习性(RR 256)和鲸目动物。Pakicetids是最早的鲸类动物,其骨盆与大多数陆地哺乳动物相似(由H-GSP 30395, 30213合成)。骨盆Ambulocetus(H-GSP 18507),但在龙王鲸(美国国家博物馆12261),骨盆不再附着在椎骨上,髂骨也大大减少。在现代弓头鲸(图中是成年雄性的骨盆)中,B.mysticetus,98B5),髋臼和闭孔丢失,髂骨减少
齿鲸目和白鲸目:现代鲸目动物
大约3400万年前,现代鲸类的第一批代表,齿类和神秘类被发现。现在一般认为齿纲动物和密纲动物(合称为新齿纲动物)起源于始新世鲸类动物的共同祖先,因此是单系的。齿齿动物身体计划中最重要的创新是获得回声定位:这些动物发出的声音被周围物体反射,这些反射使它们能够想象周围的环境。神秘菌获得了一种新颖的捕食机制:它们用嘴里的过滤器——鲸须板来过滤散装猎物(如磷虾)的饲料。尽管回声定位和滤食性分别是齿形目和密形目重要的进化主题,但这两个亚目都是多样化的,以不同的猎物为食,使用不同的狩猎技术。
齿状纲和神秘纲几乎征服了所有的海洋:它们包括沿海和近海形式、北极和热带水域、浅水、深海和河流形式。已经出版了齿类动物和神秘动物的进化史的很好的介绍(Fordyce和Muizon2001;比安努奇和兰蒂尼2007).
结论
在过去的20年里,鲸鱼的起源已经从几乎没有化石为基础,变成了宏观进化的一个最具文献记录的例子。27).尽管有这样的记录,还有更多研究的空间。例如,原生生物是多样的,其中许多鲜为人知,一些器官系统(如视觉)的进化也鲜为人知(Thewissen和Nummela)2008).

在鲸类动物进化的前1000万年中,鲸类动物身体结构的变化。图为五个古菌科,最古老的是pakicetids,而最年轻的是basilosaurids(修改自Thewissen et al.(2002),©年度审核)
尽管我们对鲸目动物起源模式的理解有所进步,但仍不清楚是哪个过程导致了这种模式:为什么鲸目动物进入海洋?丰富的新食物来源被认为是鲸类进入水中的一个原因,但这是不太可能的,因为鲸类的祖先已经生活在非常浅的淡水中。这一新发现表明,这些动物将水作为抵御危险的避难所。拉奥利德齿与早期鲸目动物的牙齿非常不同,这表明在栖息地改变后发生了饮食变化,这可能对早期鲸目动物的多样化至关重要,但对它们进入水中并没有影响。另一方面,尚不清楚海鳗吃什么,海鳗和早期鲸目动物的齿列都没有很好的现代类似物。有人认为早期鲸类动物吃鱼(O’leary和Uhen1999).
已经出现的丰富的化石记录现在可以用来丰富进化科学的其他子领域,包括发育生物学、比较解剖学和分子系统学。我们希望对进化模式的详细了解将使我们能够确定推动鲸类动物进化的过程。
参考文献
阿斯兰A, Thewissen JGM。巴基斯坦北部库尔达那组古土壤的初步评价及其对解释脊椎动物化石组合的意义。Palaeovert。1997;25:261 - 77。
Bajpai SB, Thewissen JGM。印度Harudi组和Subathu组中始新世鲸类。进:Thewissen JGM,编辑。鲸类的出现:鲸类起源的进化模式。第1版。纽约:全体会议;1998.213 - 34页。
Bajpai S, Thewissen JGM。来自Kachchh(古吉拉特邦,印度)的一种新的小型鲸鱼及其对鲸类动物运动进化的影响。Curr Sci(新德里)。2000; 79:1478 - 82。
Bajpai SB, Thewissen JGM, Kapur VV, Tiwari BN, Sahni S.印度Kachchh始新世和渐新世海牛(哺乳动物纲)。杂志,2006;26:400-10。
比安努奇G,兰蒂尼W.化石史。入职:编辑米勒·德。鲸类生殖生物学与系统发育。Enfield: Science Publ第1版;2007.35 - 94页。
脊椎形态对早期鲸类运动进化的影响。进:Thewissen JGM,编辑。鲸类的出现:鲸类起源的进化模式。第1版。纽约:全体会议;1998.325 - 52页。
Clementz MT, Goswami A, Gingerich PD, Koch PL.早期鲸鱼和海牛的同位素记录:生态过渡的对比模式。杂志,2006;26:355-70。
菲鱼。哺乳动物游泳从拖曳推进到升力推进的转变。Am Zool 1996; 36:628-41。
李志刚,李志刚。鲸类动物的进化研究进展。In: Mazin J-M, Buffrénil VD,编辑。四足动物对水中生活的二次适应。第1版。慕尼黑:弗拉格·弗里德里希·普菲尔;2001.p . 169 - 233。
盖茨西J, O'Leary MA。用分子和化石来破译鲸鱼的起源。生态学报,2001;16:562-70。
Geisler JH, Uhen MD.河马和鲸鱼之间密切关系的形态学支持。中华医学杂志,2003;23:991-6。
Geisler JH, Saunders AE, Luo Z-X。南卡罗来纳始新世中期晚期发现的一种新的原始鲸目(鲸目,古鲸目)Am Mus 2005年11月;3480:1-65。
Geisler JH, Theodor JM, Uhen MD, Foss SE。鲸目动物与陆生偶蹄动物的系统发育关系。在:Prothero DR, Foss SE,编辑。偶蹄动物的进化。巴尔的摩:约翰霍普金斯大学;2007.胜38负p。。
金格里奇PD,史密斯BH,西蒙斯EL。后肢龙王鲸伊希斯:鲸鱼有脚的证据。科学。1990;229:154-7。
Gingerich PD, Raza SM, Arif M, Anwar M, Zhou X.巴基斯坦始新世新鲸和鲸类游泳的起源。大自然。1994;368:844-7。
金格里奇PD,阿里夫M,巴蒂MA,拉扎HA,拉扎M。Protosiren而且Babiacetus巴基斯坦旁遮普Sulaiman Range始新世中Drazinda组的哺乳动物(哺乳动物,Sirenia和鲸类)。清华大学学报(自然科学版),1995a; 29:31 - 57。
金格里奇PD,阿里夫M,克莱德WC。巴基斯坦旁遮普苏莱曼山脉始新世中Domanda组的新古菌(哺乳动物,鲸目)。清华大学学报(自然科学版),1995b;29:29 - 330。
金格里奇PD,阿里夫M,巴蒂MA,安瓦尔M,桑德斯WJ。龙王鲸drazindai而且Basiloterus hussaini、新古鲸亚目(哺乳动物,鲸目)中始新世Drazinda组,以及巴基斯坦旁遮普省Sulaiman Range Khirthar群含鲸地层年龄的修正解释。清华大学学报(自然科学版)。1997;30:55-81。
Gingerich PD, Ul-Haq M, Khan IH, Zalmout I.在俾路支省苏莱曼山脉东部Drug Lahar的始新世地层和古海洋鲸(哺乳动物,鲸目)。清华大学学报(自然科学版)。2001a; 30:269-319。
Gingerich PD, Haq M, Zalmout IS, Khan IH, Malkani MS.早期偶蹄目鲸的起源:巴基斯坦始新世原生鲸科的手和脚。科学。2001 b; 293:2239-42。
北美洲中始新世原始骨化石的颅后骨学。进:Thewissen JGM,编辑。鲸类的出现:鲸类起源的进化模式。第1版。纽约:全体会议;1998.p . 235 - 67。
Hulbert RC Jr, Petkewich RM, Bishop GA, Burky D, Aleshire DP。来自格鲁吉亚的一种新的始新世中原生鲸(哺乳动物:鲸目:古鲸目)和相关生物群。张志刚,1998;
Kellogg R. Archaeoceti综述。华盛顿卡内基研究所出版,第482卷。华盛顿:华盛顿卡内基研究所;1936.1 - 366页。
卢卡斯。角龙王鲸的骨盆带(欧文),骨骼的其他部分有注释。美国自然科学1900;23:327-31。
Madar SI。早期古细菌长骨的结构适应。进:Thewissen JGM,编辑。鲸鱼的出现,鲸类起源的进化模式。第1版。纽约:全会;1998.p . 353 - 78。
Madar SI。巴基目鲸目动物的后颅骨。中国科学(d辑),2007;
Madar SI, Thewissen JGM, Hussain ST.Ambulocetus•(鲸目,陆行鲸科),以及它们对早期鲸鱼运动的影响。中华医学杂志2002;22:405-22。
Nikaido M, Rooney AP, Okada N.基于短和长穿插元素插入的鲸手目之间的系统发育关系:河马是鲸的最近的现存亲属。美国国家科学研究院。1999; 96:10261-6。
Nummela S, Thewissen JGM, Bajpai S, Hussain ST, Kumar K.鲸鱼听力的始新世进化。大自然。2004;430:776-8。
Nummela S, Hussain ST, Thewissen JGM。巴基鲸科(鲸目,哺乳目)颅骨解剖。J Vert Pal. 2006; 26:746-59。
Nummela S, Thewissen JGM, Bajpai S, Hussain ST, Kumar K.古代和现代鲸鱼的声音传播:水下听力的解剖学适应。Anat Rec. 2007; 290:716-33。
O’leary MA, Uhen MD.鲸鱼起源的时间和行为变化在陆地-水过渡中的作用。古生物学。1999;25:534-56。
Roe LJ, Thewissen JGM, Quade J, O'Neil JR, Bajpai S, Sahni A,等。用同位素方法来了解最早鲸类动物从陆地到海洋的转变。进:Thewissen JGM,编辑。鲸鱼的出现,鲸类起源的进化模式。第1版。纽约:全会;1998.p . 399 - 421。
斯波尔F,德威森JGM。水生哺乳动物平衡的比较和功能解剖。进:Thewissen JGM, Nummela S,编辑。阈值上的感觉:次生水生脊椎动物的适应。伯克利:加州大学出版社;2008.p . 257 - 86。
spor F, Bajpai S, Hussain ST, Kumar K, Thewissen JGM。早期鲸类动物水生行为进化的前庭证据。大自然。2002;417:163-6。
斯特拉瑟斯博士在大鳍鲸的原始后肢上(一道)与座头鲸和格陵兰露脊鲸相比。《物理学》,1893;27:291-335。
海鲸科(鲸目,哺乳目)的牙齿形态。中国科学(d辑),2001;
鲸鱼的起源是宏观进化的典范。BioSci。2001 b; 5:1037-49。
德威森JGM,菲什FE。最早鲸类动物的运动进化:功能模型,现代类似物和古生物证据。古生物学。1997;23:482 - 90。
德威森JGM,威廉姆斯EM.鲸目动物(鲸、海豚和鼠海豚)的早期进化。生态学报,2002;33:73-90。
水生四足动物的感觉进化:朝向和综合方法。进:Thewissen JGM, Nummela S,编辑。阈值上的感觉生物学:次生水生脊椎动物的适应。伯克利:加州大学出版社;2008.p . 333 - 330。
Thewissen JGM, Hussain ST, Arif M.考古鲸水生运动起源的化石证据。科学。1994;263:210-2。
Thewissen JGM, Madar SI, Hussain ST。Ambulocetus•来自巴基斯坦的始新世鲸类动物(哺乳纲)。中华人民共和国法律学院。1996;190:1-86。
Thewissen JGM, Williams EM, Hussain ST.印度-巴基斯坦北部始新世哺乳动物动物群。J Vert Pal. 2001; 21:347-66。
Thewissen JGM, Cohn MJ, Stevens LS, Bajpai S, Heyning J, Horton WE Jr.海豚后肢丧失的发育基础和鲸类身体计划的起源。美国国家科学研究院。2006; 103:8414-8。
Thewissen JGM, Cooper LN, Clementz MT, Bajpai S, Tiwari BN。鲸起源于印度始新世时期的水生偶蹄动物。大自然。2007;450:1190-5。
始新世中晚期龙王蜥。进:Thewissen JGM,编辑。鲸类的出现:鲸类起源的进化模式。第1版。纽约:全体会议;1998.29 - 61页。
的形态、功能和解剖Dorudon atrox(哺乳动物,鲸目):埃及始新世中晚期的一种古动物。大学学报(自然科学版)2004;34:1-222。
最早的鲸目动物概要:巴基鲸科,陆行鲸科,雷明鲸科和原鲸科。进:Thewissen JGM,编辑。鲸类的出现:鲸类起源的进化模式。第1版。纽约:全体会议;1998.28页。
确认
我们感谢巴基斯坦地质调查局在收集和研究巴基斯坦化石方面的合作以及后勤支持,感谢S. Taseer Hussain博士领导霍华德大学-巴基斯坦地质调查局项目。我们感谢阿贾伊·塔科尔和古吉拉特邦矿业开发公司在古吉拉特邦的实地工作方面提供的援助,感谢布吉的巴蒂先生在后勤方面提供的帮助。我们感谢阿拉斯加爱斯基摩捕鲸委员会和巴罗捕鲸船长协会提供标本并为他们的科学研究做出贡献。弓头鲸标本是根据NMFS海洋哺乳动物采集许可证814-1899收集的。我们还要感谢北坡区野生动物部和巴罗北极科学联盟在标本采集方面的后勤支持和协助。这项工作得到了印度科技部(给Sunil Bajpai)和美国国家科学基金会(给j.g.m. Thewissen)的资助。
作者信息
作者及隶属关系
相应的作者
权利和权限
开放获取本文根据创作共用属性2.0国际许可协议(https://creativecommons.org/licenses/by/2.0),允许在任何媒介上不受限制地使用、传播和复制,前提是正确地引用原始作品。
关于本文
引用本文
德威森,j.g.m.,库珀,l.n.,乔治,J.C.et al。从陆地到水:鲸、海豚和鼠海豚的起源。Evo Edu Outreach2, 272-288(2009)。https://doi.org/10.1007/s12052-009-0135-2
收到了:
接受:
发表:
发行日期:
DOI:https://doi.org/10.1007/s12052-009-0135-2
关键字
- 海豚
- 鲸鱼
- 海豚
- 进化
- 鲸类
- 哺乳动物