摘要
背景
冷诱导rna结合蛋白(CIRP)是低温治疗的关键蛋白。高表达的CIRP对神经元具有神经保护作用。本研究旨在为CIRP对神经胶质细胞的保护作用提供证据,并探索CIRP的下游通路。
结果
本研究结果表明,与对照组相比,CIRP过表达的胶质细胞的细胞活力显著增加。CIRP过表达后,表皮生长因子(EGF) mRNA表达量显著升高,衍生性神经营养因子(BDNF)、bcl-2、血管内皮生长因子(VEGF)和神经生长因子(NGF) mRNA表达量较对照组无上调。EGF和CIRP在胶质细胞上共表达。与对照组相比,表达CIRP后,胶质细胞上EGF的表达显著增加。
结论
CIRP过表达可提高神经胶质细胞的活力,发挥神经保护作用。在CIRP过表达的神经胶质细胞上,EGF的表达被激活,这暗示了通过EGF激活CIRP神经保护的途径。
背景
冷诱导RNA结合蛋白(CIRP)是一种由n端RNA识别基序和c端富含精氨酸区组成的18kd蛋白。它主要定位于细胞核,可以迁移到结合mRNA的细胞质中。冷应激下脑内CIRP表达显著增加[1,2],而CIRP的上调在触发动物冬眠中起着重要作用[3.,4].这些发现激发了高表达CIRP可能发挥神经保护功能的想法。我们之前的研究表明CIRP可以抑制神经元凋亡[5,6], Zhang等[7]报道缺氧条件下过表达的CIRP可以恢复神经干细胞的增殖,提示CIRP具有神经保护作用。最近一些研究表明,胞外CIRP可诱导炎症反应,导致细胞损伤[8],那么直接使用CIRP作为神经保护因子在理论上似乎是不合理的。我们假设,当CIRP在细胞内高表达而细胞外分布最小时,这是作为神经保护因子的理想条件。细胞内CIRP表达的通路可能为实用的神经保护方案提供新的线索。我们之前的工作只提出CIRP的增加可以通过增加Bcl-2、Akt和pERK1/2蛋白来减少神经元凋亡[5].越来越多的研究强调神经胶质细胞在神经保护功能中的作用。在本研究中,我们研究了CIRP过表达对神经胶质细胞系细胞活力的影响,以及CIRP表达与一些神经保护因子,包括表皮生长因子(EGF)、衍生神经营养因子(BDNF)、bcl-2、血管内皮生长因子(VEGF)和神经生长因子(NGF)的关系。
材料与方法
细胞培养
胶质细胞系SHG-44购自上海GeneChem有限公司(上海,中国)。细胞培养于96孔板中,密度约为每孔10,000个细胞,培养基为GibcoDulbecco 's Modified Eagle Media (DMEM/F-12;赛默飞世尔科技公司),含10%胎牛血清(FBS,赛默飞世尔科技公司),补充1%青霉素/链霉素(赛默飞世尔科技公司),在37℃,5% CO的湿化气氛中2在空中。在收集细胞前24 h,每周更换培养基2-3次。
CIRP在SHG-44细胞过表达的建立
本研究使用的含绿色荧光蛋白(GFP)报告基因的重组慢病毒载体系统购自上海Genechem有限公司。构建了编码CIRP基因AB000362全长的载体,通过PCR和DNA测序证实了CIRP cDNA的准确插入。转染时,将胶质细胞置于96孔板中,用CIRP慢病毒载体(CIRP过表达组,e -CIRP)或GFP慢病毒载体(阴性GFP对照组,N-GFP)在无血清培养基中转染12 h,感染复数(MOI)为40。加入5ug/mL的聚苯乙烯,提高感染效率。其余未感染慢病毒的细胞作为对照组(control group, CON),转染naïve慢病毒的细胞作为模拟组(mock group, mock)。然后将细胞洗涤并包埋到完整的培养基中。转染4天后,荧光显微镜检测GFP基因表达。计算转染效率。
细胞治疗
OE-CIRP细胞和模拟细胞分别遭受氧化、缺氧、葡萄糖剥夺和谷氨酸诱导的神经毒性等4种损伤。首先,用不同浓度的H处理模拟细胞2O2, CoCl2和谷氨酸分别作用12小时,选择合适的浓度进行下一步操作。选定浓度确定后,进行4次损伤。H2O2如下。先除去介质,再除去H2O2被添加到细胞中。培养基立即加入到孔中,得到最终选定的H浓度2O2。每2小时重复此过程,直到12小时进一步分析。CoCl2(232,696, Sigma-Aldrich)在选定的浓度下制备成水溶液,诱导化学缺氧。用CoCl处理神经胶质细胞2用不含葡萄糖和无血清的DMEM (11,966,025, Thermo Fisher Scientific)培养细胞12小时,以达到葡萄糖剥夺。将细胞以选定浓度的谷氨酸暴露12小时以诱导神经毒性。
MTT法测定细胞活力
在进行MTT检测前,将培养基替换为100µL新鲜培养基。每孔加入MTT溶液(10µL, 12 mM, Vybrant™MTT Cell Proliferation Assay Kit, Thermo Fisher Scientific),包括10µL MTT溶液单独加入100µL培养基的阴性对照。37°C孵育4小时后,用微孔板仪(Fluoroskan Ascent, Thermo Fisher Scientific)分析细胞活力,吸光度为570 nm。
实时聚合酶链反应
分离总RNA,合成cDNA,如我们之前报道的[1].本研究使用的引物组合由中国上海Sangon biotech设计并提供,列于表中1。实时逆转录酶(RT-PCR)分析使用SLAN实时荧光定量PCR系统(上海洪石,中国)。扩增所用的PCR条件为:95℃初始变性5 min, 95℃变性15 s 40次循环,65℃退火30 s, 84℃延伸30 s。在每个周期结束时实时收集数据。每个反应包括阴性对照,不含逆转录酶和水对照。除了对反应进行实时分析和熔化曲线分析外,扩增产物在2%琼脂糖凝胶上与溴化乙啶进行电泳分离,并在紫外光下观察,以确认扩增子大小合适,并且没有非特异性产物。所有PCR产物均产生单一特异性产物。
相对表达数据按二δ阈值(∆∆Ct)相对定量法计算。通过从每个样本获得的CIRP Ct值中减去β-actin Ct来进行Ct值的归一化。为了计算样本之间循环次数的相对差异,选择对照组样本的平均Ct值作为基线。这个数字是从之前归一化为β-肌动蛋白的每个Ct值中减去的。采用2-Δ(ΔCt)计算各组间Ct值的相对差异。
双免疫组织化学
将细胞用PBS孵育2次,每次10分钟,然后用4%多聚甲醛在室温下固定15分钟,然后用含0.05% Tween20的PBS洗涤以去除固定物。将固定细胞在室温下用0.1% Triton X-100在TBS中渗透10分钟,在室温下用1%阻断剂BSA阻断15分钟。抗cirp抗体(AB166775, Abcam)和抗egf抗体(701538,Invitrogen)作为一抗分别加入相应的孔中,在4°C下孵育过夜。一抗孵育后,用PBS洗涤细胞,然后与Alexa Fluor 488驴抗山羊IgG二抗和Alexa Fluor Plus 594驴抗兔IgG二抗室温孵育2小时。用PBS洗涤细胞,然后用DAPI核染色(D9542,默克)孵育1分钟。图像是在奥林巴斯共聚焦显微镜上拍摄的。
免疫印迹
用冷PBS (0.01 M pH7.2~7.3)洗涤细胞2次,用1mM PMSF (8553, cell Signaling Technology)重悬于冷冻细胞裂解缓冲液(9803,cell Signaling Technology)中,4°C孵育30 min。裂解物在4°C下以12,000 rpm离心10 min。上清蛋白保存于- 80℃。用CIRP单克隆抗体(AB166775, Abcam)和EGF单克隆抗体(701538,Invitrogen)进行Western blotting分析CIRP和EGF水平。以β-肌动蛋白为内对照。使用光学扫描仪和Labworks系统(Labworks Inc ., Winnipeg, Manitoba, Canada)对印迹条带进行定量。
统计分析
Windows采用Sigma Plot(12.5)进行统计分析。数据以三个独立实验的平均值±标准差表示。差异的统计分析采用单因素方差分析。P以< 0.05为有统计学意义。
结果
细胞转染及CIRP在胶质细胞上的过表达
培养细胞在光镜下呈现胶质细胞的特性(图2)。1B, D, F)。用GFP或CIRP慢病毒载体转染4天后,在荧光显微镜下观察GFP表达的细胞(图2)。1C, E, G)。MOI为40时,感染效率在80%以上。RT-PCR分析显示,与GFP对照组(1.07±0.12)相比,CIRP组CIRP mRNA的相对水平(5476.56±442.8)显著升高(图2)。1一个)。

CIRP mRNA在神经胶质细胞系SHG-44中的过表达。一个正常对照细胞(CON)间CIRP mRNA表达的比较(n= 3),阴性对照细胞转染绿色荧光蛋白(N-GFP) (n= 3),转染了CIRP的细胞(OE-CIRP) (n= 3) *P< 0.01。B光镜下正常SHG-44细胞(×200)。(C)荧光显微镜下正常SHG-44细胞(×200)。D光镜下转染GFP的SHG-44细胞(×200)。E荧光显微镜下转染GFP的SHG-44细胞(×200)。F光镜下转染CIRP的SHG-44细胞(×200)。G荧光显微镜下转染CIRP的SHG-44细胞(×200)
CIRP过表达的神经胶质细胞的细胞活力
模拟细胞的细胞活力随着H浓度的升高而降低2O2, CoCl2谷氨酸增加(图2)。2a - c)。选择细胞活力在50%左右的浓度作为适宜浓度。神经胶质细胞暴露于50µmol/L H的氧化作用后2O212小时后,与对照组相比,过表达的CIRP显著提高了细胞活力(77.6±4.9% vs 39.4±2.8%)。2D). 75µmol/L CoCl诱导12 h化学缺氧后2与对照细胞相比,过表达的CIRP也显著提高了胶质细胞的细胞活力(74.7±4.3%比39.4±5.1%)(图2)。2经过12小时的葡萄糖剥夺处理,与对照组相比,过表达的CIRP显著提高了细胞活力(64.8±2.1%对33.2±4.2%)(图2)。25µmol/L谷氨酸处理12 h后,与对照细胞相比,过表达的CIRP细胞活力显著提高(73.2±5.2% vs 50.3±7.1%)(图2)。2D)。
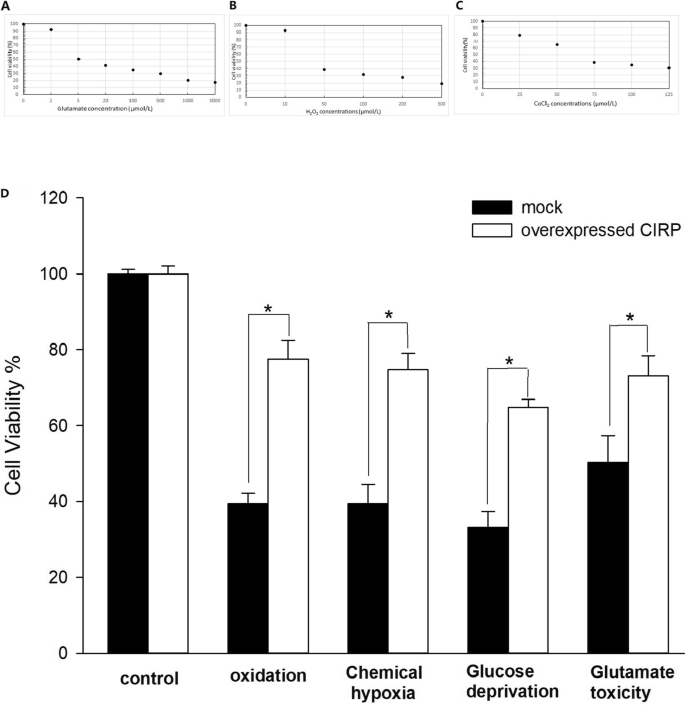
不同浓度的H2O2(一个), CoCl2(B)和谷氨酸(C)在神经胶质细胞上。氧化、缺氧、葡萄糖剥夺和谷氨酸中毒处理后的神经胶质细胞活力(D).实验重复了三次。条形图表示平均值±SD。* p < 0.05
CIRP过表达后生长因子和bcl-2在胶质细胞上的表达
分析了几种具有神经保护作用的生长因子和bcl-2在CIRP过表达的胶质细胞中的表达。与阴性对照相比,CIRP过表达的细胞中EGF mRNA的相对表达量显著增加(图2)。3.A)。与阴性对照相比,CIRP过表达的细胞中BDNF mRNA的相对表达量有统计学意义上的降低(图2)。3.B)。bcl-2与bcl-2相对mRNA表达量差异无统计学意义(图2)。3.C), VEGF(无花果。3.D)、神经生长因子(无花果。3.E)和阴性对照组。
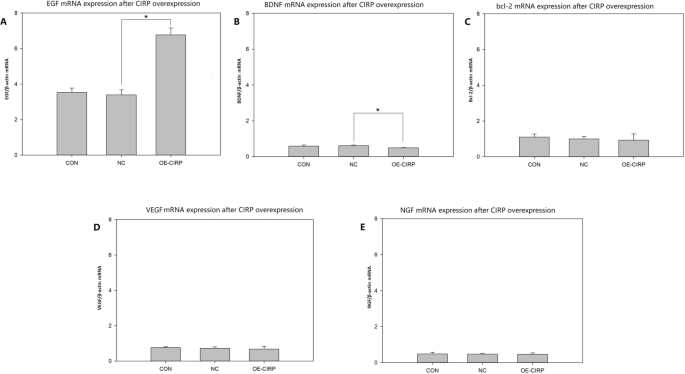
RT-PCR分析CIRP在神经胶质细胞系SHG-44上过表达后部分神经营养因子和细胞因子的表达。一个EGF mRNA表达量(n= 3), *p< 0.05。BBDNF mRNA表达量(n = 3), *p< 0.05。Cbcl-2 mRNA表达(n = 3)。DVEGF mRNA表达量(n= 3)。FNGF mRNA表达量(n= 3) CON:正常对照细胞组;NC:阴性对照细胞组,转染阴性对照慢病毒载体的细胞;OE-CIRP:过表达CIRP细胞组,转染CIRP慢病毒载体的细胞
CIRP过表达增加了胶质细胞上EGF的表达
用CIRP慢病毒载体转染4天后,对胶质细胞进行CIRP和EGF的双重免疫组化。双染色显示CIRP在胶质细胞上强烈表达(图2)。4B),同时,EGF也在胶质细胞上显著表达(图2)。4A)。在合并图像中,CIRP和EGF在胶质细胞上的分布几乎重合,CIRP主要在细胞核内,EGF在细胞核周围呈点状分布(图2)。4D)。对这些细胞进行western blot分析显示,与阴性对照组相比,过表达组的CIRP表达显著增加(0.85±0.1%比0.12±0.06%)。4同时,与阴性对照组相比,CIRP过表达组的EGF表达也显著增加(0.52±0.01%比0.21±0.09%)(图2)。4E)。

CIRP过表达增加了胶质细胞的EGF水平。双标记免疫荧光染色和共聚焦显微镜观察EGF(绿色)一个), CIRP(红色)B), DAPI染色(蓝色C)和合并(D).Western blot分析CIRP和EGF,以及CIRP和EGF的相对表达E。CON:正常对照组,MOCK:阴性对照组,OE: CIRP过表达组。实验重复了三次。条形图表示平均值±SD。* p < 0.01
讨论
大多数临床前研究表明,诱导治疗性低温对不同的脑损伤具有神经保护作用。在临床实践中,低温治疗通常被推荐作为脑损伤和缺血性脑卒中的治疗方法。然而,治疗严重创伤性头部损伤的最新指引[9]或急性缺血性中风[10]仅将治疗性低温列为BII推荐,这意味着证据质量较低。治疗性低温在临床应用时的局限性主要在于体温下降时的躯体颤抖、意识改变等并发症。“目标温度管理”的概念[11,12]是近年来兴起的,试图克服这些局限[13,14,15],但结果仍不能满足临床实践的需要。精确控制大脑温度似乎是达到神经保护效果的最好方法,同时避免低体温引起的并发症。CIRP现在被认为是冷应激的可能启动子和“目标温度管理”的理想操纵靶标。但CIRP的生物活化机制和下游通路仍有待阐明。
在分子水平上,CIRP是治疗操作的潜在靶点,以达到低脑温度的效果。由于胞外CIRP可引起严重的炎症反应,损伤细胞[8],应首先验证高表达CIRP在细胞上的安全性。我们的研究表明,细胞内过表达的CIRP在形态学上不影响胶质细胞的生长(图2)。1).此外,CIRP过表达的神经胶质细胞在氧化、缺氧、葡萄糖剥夺和谷氨酸毒性等不同有害条件下的活力均有所增加(图2)。2).我们前期的研究已经证明了CIRP对脑神经元有保护作用2O2诱导凋亡[5,6].本研究结果为CIRP对神经胶质细胞具有明显的神经保护作用提供了直接证据。近年来,神经胶质细胞在中枢神经系统(CNS)中的作用越来越受到关注。脑内胶质细胞被认为是调节神经元功能的重要组成部分,除了维持神经元网络和内稳态外[16,17,18].Toriuchi K.等[19在低温条件下,促红细胞生成素表达增加的星形胶质细胞对神经元具有神经保护作用。新兴研究认为神经胶质细胞是脑疾病和损伤治疗药物的有吸引力的靶点[18].本研究支持了调控神经胶质细胞的CIRP是保护不同损伤后脑功能的潜在途径。细胞内CIRP过表达可延长细胞周期G1期,抑制细胞生长速率,从而降低细胞的能量需求[20.].这一机制可以部分解释CIRP过表达的胶质细胞的细胞活力增加。
目前,人们普遍认为冷应激、紫外线照射和一定程度的缺氧可诱导不同细胞上CIRP的表达[21].但是这些物理压力如何影响CIRP表达的确切机制仍然未知,并且操纵CIRP过表达对细胞的影响也需要阐明。以往的研究[22表明CIRP是在寒冷环境中表达增加的有限基因之一。CIRP可能是激活下游通路以适应低温的启动子。自2006年以来,我们对CIRP进行了研究,但除了慢病毒转染外,我们没有发现任何有效的生物激活CIRP表达的方法。CIRP的下游通路也是实现治疗性低温的合理操纵点。首先,我们想知道CIRP是否可以触发任何已知的神经保护因子和细胞因子特异性地发挥其功能。我们分析了EGF、VEGF、NGF、BDNF和bcl-2基因在CIRP过表达的胶质细胞中的表达情况。结果显示,只有EGF mRNA表达和EGF表达显著升高(图2)。3.和4).EGF是一个被广泛研究的因子,它通过激活EGFR家族在促进细胞增殖和对抗细胞凋亡中发挥关键作用。EGF以神经营养的方式作用于中枢神经系统的几个成熟神经元,增加神经元的存活和生长过程,防止神经元因神经毒性、氧化和其他有害条件而死亡[23].有趣的是,EGF对神经元的神经保护作用不仅来自于对神经元的直接作用,还来自于通过胶质细胞的细胞间作用[23].本研究在常温下细胞内CIRP过表达的情况下,EGF mRNA和EGF表达均显著升高,且CIRP和EGF在胶质细胞中的分布几乎重合(图2)。4D)。一些文献报道,在冷胁迫下,细胞中只有CIRP、RBM3和NF-1变体的表达增加[22].然后我们假设在复温时,激活的CIRP在细胞质中迁移,并可以立即触发EGF的表达。该通路可能对神经胶质细胞和神经元具有重要的保护作用。Kaija H.等。[24]报道,健康大鼠前列腺组织在低温复温过程后,双调节蛋白(AMR)、细胞周期蛋白D1 (CyD1)、细胞周期蛋白依赖性激酶抑制剂1a (p21)、跨膜形式前列腺酸性磷酸酶(PAcP)、血栓调节蛋白(TM)、热休克转录因子1 (HSF1)和Bax/Bcl-2的相对mRNA表达量显著升高。目前尚不清楚这种模拟临床低温治疗的低温复温过程如何影响神经元和神经胶质细胞的基因表达。EGF现已成为细胞培养领域中促进细胞分化、成熟和存活不可缺少的试剂。但如果没有其他细胞因子和因子的协同作用,单个EGF无法完成这一复杂的任务。需要进一步的研究来完全了解CIRP激活的下游途径。
结论
本研究表明,通过转染CIRP慢病毒载体,CIRP mRNA在胶质细胞中的表达显著增加。CIRP在神经胶质细胞中的过表达增加了细胞在氧化、化学缺氧、葡萄糖剥夺和谷氨酸神经毒性下的活力,而CIRP的过表达也增加了神经胶质细胞中EGF mRNA和EGF的表达,这表明在CIRP激活后,EGF可能作为神经保护因子发挥关键作用。
数据和材料的可用性
支持本研究结果的数据可根据合理要求从通讯作者AL处获得。
参考文献
刘安,张志,李安,薛杰。低温和脑缺血对大鼠脑冷诱导rna结合蛋白mRNA表达的影响。中国生物医学工程学报,2010;17(1):1 - 4。
薛建辉,Nonoguch K, Fukumoto M, Sato T, Nishiyama H, Higashitsuji H,等。缺血和h2o2对大鼠神经细胞冷应激蛋白cirp表达的影响。中华放射医学杂志。1999;27(11/12):1238-44。
Shiina T, Shimizu Y.前体mrna的温度依赖性选择性剪接及其生物学意义:冬眠哺乳动物冷休克蛋白基因转录后调控的综述。中国生物医学工程学报,2011;31(2):444 - 444。
Logan SM, Storey KB。冷诱导rna结合蛋白cirp,而非Rbm3,可能调控冬眠地鼠组织中转录产物的加工和保护。细胞应激伴侣。2020;25(6):857-68。
张海涛,薛建华,张志伟,孔华兵,刘爱军,李世昌,等。冷诱导rna结合蛋白通过抑制线粒体凋亡抑制神经元凋亡。中国生物医学工程学报,2015;22(2):474 - 483。
李松,张志,薛军,刘安,张慧。冷诱导RNA结合蛋白抑制H(2)O(2)诱导的大鼠皮层神经元凋亡。中国生物医学工程学报,2012;41(1):447 - 452。
张强,王亚忠,张伟,陈旭,王健,陈健,等。冷诱导rna结合蛋白参与体外重度缺氧诱导的神经干细胞生长停滞。中国生物医学工程学报,2017;35(3):444 - 444。
王鹏,王平。细胞外CIRP (eCIRP)与炎症的关系。中国生物医学工程学报,2019;31(1):444 - 444。
Carney N, Totten AM, O 'Reilly C, Ullman JS, Hawryluk GW, Bell MJ,等。严重创伤性脑损伤管理指南,第四版。神经外科。2017;80(1):6 - 15。
华纳JJ,哈灵顿RA,萨科RL,埃尔金德MSV。急性缺血性卒中患者早期管理指南:2019年对2018年急性缺血性卒中早期管理指南的更新2019; 50(12): 3331 - 2。
Nielsen N, weterslev J, Cronberg T, Erlinge D, Gasche Y, Hassager C,等。目标温度控制在33摄氏度和心脏骤停后的36摄氏度。中华医学杂志,2013;39(3):397 - 396。
Jackson TC, Kochanek PM。靶向温度管理时代治疗性低温的新愿景:一种推测性综合。治疗性低温和体温管理。2019; 9(1): 13-47。
Jain A, Gray M, Slisz S, Haymore J, Badjatia N, Kulstad E.寒战治疗的针对性温度管理研究进展。中华神经科学杂志,2018;50(2):63-7。
Dietrich WD, Bramlett HM。创伤性脑损伤的治疗性低温和目标温度管理:成功翻译的临床挑战。中国生物医学工程学报,2016;32(1):444 - 444。
Madden LK, Hill M, May TL, Human T, Guanci MM, Jacobi J,等。目标体温管理的实施:来自神经危重症护理学会的循证指南。神经内科杂志,2017;27(3):468-87。
Allen NJ, Lyons DA。神经胶质是中枢神经系统形成和功能的建筑师。科学。2018;362(6411):181 - 5。
Nakano M, Tamura Y, Yamato M, Kume S, Eguchi A, Takata K,等。NG2胶质细胞调节神经免疫反应,维持神经元功能和存活。科学通报,2017;7:442 - 441。
王晓明,王晓明。星形胶质细胞在创伤性脑损伤中的病理生理反应。中华生物医学杂志,2011;22(12)。
田村,李建平,李建平,李建平。低温条件下星形胶质细胞衍生的促红细胞生成素的表达对神经损伤的影响。中华神经科杂志,2010;17(1):1 - 4。
李建军,李建军,李建军,李建军,李建军,等。一种富含甘氨酸的rna结合蛋白对动物细胞生长的抑制作用。中国生物医学工程学报,2009;37(4):391 - 391。
钟萍,黄华。冷诱导rna结合蛋白的研究进展。未来科学OA。3 (4): FSO246-FSO。2017;
Sonna LA, Fujita J, Gaffin SL, Lilly CM。特邀综述:热应激和冷应激对哺乳动物基因表达的影响。应用生理学杂志(Bethesda, Md: 1985)。2002年,92(4):1725 - 42。
王晓明,王晓明,王晓明,等。表皮生长因子对神经营养的影响。中国生物医学工程学报,2009;31(1):391 - 391。
李建军,李建军,李建军,等。低温和复温对正常大鼠前列腺组织基因表达和细胞增殖的影响。PLoS ONE。2015; 10 (5): e0127854。
致谢
(不适用)
资金
本研究得到了中国科技部国际科技合作计划(2011DFA30550)的资助。
作者信息
作者及单位
贡献
AL和ZZ是主要研究者并撰写了这份手稿文本,SL, HZ, YJ和HK进行了研究并准备了草稿。所有作者都审阅并批准了手稿的最终版本。
相应的作者
道德声明
伦理批准并同意参与
(不适用)
发表同意书
(不适用)
相互竞争的利益
作者声明不存在经济或其他方面的利益冲突。
额外的信息
出版商的注意
伟德体育在线施普林格·自然对已出版的地图和机构关系中的管辖权要求保持中立。
权利和权限
开放获取本文遵循知识共享署名4.0国际许可协议,该协议允许以任何媒介或格式使用、共享、改编、分发和复制,只要您适当地注明原作者和来源,提供知识共享许可协议的链接,并注明是否进行了更改。本文中的图像或其他第三方材料包含在文章的知识共享许可协议中,除非在材料的署名中另有说明。如果材料未包含在文章的知识共享许可中,并且您的预期用途不被法律法规允许或超过允许的用途,您将需要直接获得版权所有者的许可。如欲查阅本许可证副本,请浏览http://creativecommons.org/licenses/by/4.0/。创作共用公共领域免责声明(http://creativecommons.org/publicdomain/zero/1.0/)适用于本文中提供的数据,除非在数据的信用额度中另有说明。
关于本文
引用本文
刘爱安,李世生,焦勇。et al。过表达冷诱导rna结合蛋白可提高胶质细胞的细胞活力和EGF的表达。BMC Mol与细胞生物学23, 58(2022)。https://doi.org/10.1186/s12860-022-00460-3
收到了:
接受:
发表:
DOI:https://doi.org/10.1186/s12860-022-00460-3
关键字
- Cold-inducibleRNA-binding蛋白质
- 神经保护
- 表皮生长因子
- 细胞生存能力